49 Congreso Internacional del Americanistas (ICA) |
|
Quito Ecuador7-11 julio 1997 |
|
 Congresos en cdrom 2.2 - version en línea
Congresos en cdrom 2.2 - version en línea
Integridad del Registro Faunístico en el Sudeste de Tierra del Fuego, Argentina
José Luis Lanata*
Introducción
Los análisis faunísticos han experimentado un importante desarrollo durante los últimos 20 años. En líneas generales han mostrado su utilidad en la discusión de distintos aspectos económicos relacionados con la explotación de estos recursos. Aquí destacamos el aporte que el registro faunístico ha realizado al conocimiento de la integridad del registro arqueológico.
Tomamos el caso de Península Mitre, Tierra del Fuego (Argentina) y estudiaremos la integridad de las muestras zooarqueológicas recuperadas por el Proyecto Arqueológico Norte de Península Mitre (1984-1991). Nuestro interés es ver si existe o no variabilidad en la integridad tanto en el nivel de las diferentes especies faunísticas dentro de cada una de las muestras como en el de su procedencia ambiental. Utilizaremos diferentes herramientas analíticas para acercarnos a esta problemática, pues entendemos que no existe una única manera de hacerlo. Con el fin de conocer el grado de integridad y calibrar las muestras, nos centraremos en el estudio de 33 sub-conjuntos faunísticos recuperados en 11 sitios. Nuestras categorías de análisis serán los restos óseos de pinnípedos, aves y guanacos (Lama guanicoe), ya que conforman genéricamente los recursos faunísticos más explotados en la región. En el caso de los pinnípedos los tratamos globalmente, al igual que las aves, donde agrupamos las diferentes especies y géneros reconocidas en las muestras. En el Apéndice adjunto presentamos las diferentes tablas con los datos utilizados aquí.
Las muestras
Las muestras tratadas en este trabajo provienen de tres localidades del sudeste de la Isla Grande de Tierra del Fuego, Argentina; región conocida como Península Mitre. De norte a sur, las mismas son Ea. María Luisa (66°47' S, 54°50' W), Rancho Donata (65°54 S, 54°66' W) y Bahía Thetis (65°26' S, 54°66' W), encontrándose las dos primeras en el ambiente denominado parque fueguino y la restante en el bosque cordillerano (Bondel 1988 ). Sin embargo es importante destacar que tanto Rancho Donata como Ea. María Luisa se encuentran situadas en diferentes sectores del parque fueguino. La primera localidad esta localizada en una amplia pradera en tanto que la segunda lo está en pleno bosque antiguo. La Tabla 1 resume las principales propiedades estructurales de los sitios de donde provienen las muestras, conjuntamente con los fechados disponibles.
|
muestra |
mo |
ma |
co |
pop |
pox |
pom |
ta |
ex |
lo |
fechado* |
|
|
Ea. |
ML5 |
montic |
valvas |
si |
- |
83 |
4 |
60 |
6 |
7 |
360 ± 50 |
|
María |
ML7 |
chato |
valvas |
si |
13 |
18 |
5 |
60 |
6 |
7 |
690 ± 50 |
|
Luisa |
MLA3 |
chato |
valvas |
si |
14 |
22 |
4 |
117 |
12 |
9 |
1020 ± 80 |
|
MLB5 |
chato |
valvas |
si |
10 |
12 |
6 |
81 |
12 |
3 |
1110 ± 60 |
|
|
Rancho |
RD"A" |
chato |
arena |
no |
31 |
43 |
29 |
175 |
7 |
1 |
1380 ± 70 |
|
Donata |
CM 1 |
chato |
valvas |
no |
22 |
25 |
18 |
36 |
3 |
3 |
- - |
|
RD 7 |
montic |
valvas |
no |
44 |
89 |
0 |
520 |
260 |
1 |
1500 ± 50 |
|
|
CTS2 |
chato |
valvas |
si |
45 |
33 |
62 |
36 |
4 |
7 |
230 ±50 |
|
|
Bahía |
Aleph 1 |
montic |
valvas |
si |
16 |
11 |
24 |
140 |
2 |
3 |
330 ± 50 |
|
Thetis |
Aleph 2 |
chato |
valvas |
si |
9 |
8 |
12 |
56 |
1 |
3 |
- - |
|
Aleph 3 |
chato |
valvas |
no |
nd |
nd |
nd |
21#>/p> |
1.5 |
3 |
450 ± 60 |
Tabla 1. Propiedades estructurales de los sitios analizados.
Ref.: mo = morfología del sitio, ma = matriz, co = columna de moluscos, pop = potencia promedio (cm), pox = potencia máxima (cm), pom = potencia mínima (cm), ta = tamaño (m²), ex = excavado (m²), lo = localización ambiental y exposición (Kvamme 1985 :218), * fechados: todos expresados en años radiocarbónicos BP. #: tamaño estimado.
Si bien restos de moluscos, roedores, cetáceos y cánidos se han registrado en algunas de la muestras, nuestro análisis se centrará exclusivamente en los restos de guanaco (G), aves (A) y pinnípedos (P). No se discutirán aspectos económicos ya que ha sido motivo de otros trabajos [1] . Tan solo analizaremos aspectos que tengan que ver con la integridad de cada una de ellas. Con este fin analizaremos aspectos relacionados con las relaciones entre los diferentes índices de cuantificación, huellas y marcas y densidad ósea. Pasamos a continuación a detallar cada uno de los análisis.
La Relación entre NISP, MNE y MNI en las Muestras
La exploración de las relaciones entre los diferentes índices de cuantificación de restos faunísticos es un punto importante para entender el grado de integridad de las muestras bajo estudio (Grayson 1978 y 1984 , Binford 1981 y 1984 , Klein y Cruz-Uribe 1984 , Mengoni 1988 , Lyman 1994 ).
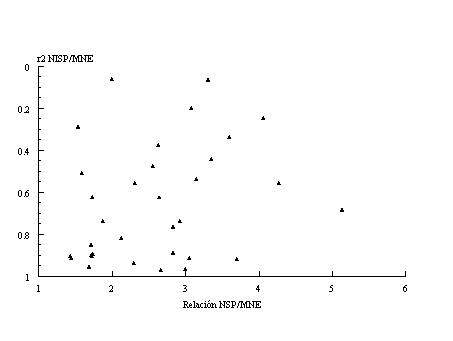
La Figura 1 presenta la relación entre NISP y MNE -este último calculado según Binford 1984. La simple lectura de la figura muestra la estrecha relación entre ambos índices en todas las muestras [2] .
|
|
Figura 1. Relación entre NISP y MNE de los sub-conjuntos faunísticos del Norte de Península Mitre. >/p> Datos procedentes de Apéndice, Tablas A1 a A6. |
La relación entre NISP y MNE ha sido utilizada como un indicador de grado de fragmentación de los conjuntos (Richardson 1980, Lanata et al. 1992 , Savanti 1994 ). En este trabajo utilizaremos los valores de r² entre ambas medidas [3] (Lanata et al. 1992). Cuando los valores de r² son más cercanos a 1 están indicando que los conjuntos se encuentran poco fragmentados -en forma independiente de la/s causa/s- incrementándose la fragmentación a medida que los valores se reducen. Esto no indica de ninguna manera los posibles agentes y/o procesos involucrados; tan solo intenta comparar el grado de fragmentación entre muestras.
Nuestra expectativa general es que los sub-conjuntos de aves, pinnípedos y guanaco se comporten diferencialmente en relación a esta propiedad del registro arqueofaunístico. Las aves serían el sub-conjunto que presente un bajo estado general de fragmentación, mientras que para el guanaco esperamos una alta fragmentación. Por su parte, una fragmentación media sería la característica para los pinnípedos
En la Tabla 2a rankeamos los valores de r² NISP/MNE de los distintos sub-conjuntos analizados, separados en pinnípedos (P), guanaco (G) y aves (A). Puede observarse -Tabla 3- que de las 33 muestras, 14 presentan valores altos -entre 1 y .800.
|
Sub-Conjunto |
r² NISP/MNE |
Ranking |
Sub-Conjunto |
relación NSP/MNE |
Ranking |
|||||
|
CTS 2 G |
0.969 |
|
MLA3 G |
5.136 |
|
|||||
|
ALEPH 1 G |
0.967 |
|
MLB5 G |
4.275 |
|
|||||
|
CTS 2 A |
0.958 |
|
ML5 G |
4.066 |
|
|||||
|
RD "A" A |
0.934 |
|
ML5 P |
3.702 |
|
|||||
|
ML5 P |
0.917 |
|
RD "A" P |
3.6 |
|
|||||
|
ALEPH 3 A |
0.915 |
|
ML7 G |
3.357 |
|
|||||
|
RD "A" G |
0.914 |
|
ML7 P |
3.312 |
|
|||||
|
RD 7 P |
0.902 |
|
MLA3 P |
3.150 |
|
|||||
|
CM1 A |
0.902 |
|
ALEPH 1 P |
3.087 |
|
|||||
|
ALEPH 1 A |
0.897 |
|
RD "A" G |
3.055 |
|
|||||
|
RD7 A |
0.896 |
|
ALEPH 1 G |
3 |
|
|||||
|
ALEPH 3 G |
0.889 |
|
ALEPH 2 P |
2.923 |
|
|||||
|
MLA3 A |
0.852 |
|
ALEPH 3 G |
2.833 |
|
|||||
|
ML5 A |
0.816 |
|
ALEPH 2 G |
2.833 |
|
|||||
|
ALEPH 2 G |
0.765 |
|
CTS 2 G |
2.666 |
|
|||||
|
ALEPH 2 P |
0.739 |
|
CM1 P |
2.642 |
|
|||||
|
CM1 G |
0.735 |
|
CTS 2 P |
2.636 |
|
|||||
|
MLA3 G |
0.685 |
|
ALEPH 3 P |
2.562 |
|
|||||
|
CM1 P |
0.624 |
|
MLB5 A |
2.314 |
|
|||||
|
ML7 A |
0.624 |
|
RD "A" A |
2.305 |
|
|||||
|
MLB5 G |
0.555 |
|
ML5 A |
2.132 |
|
|||||
|
MLB5 A |
0.554 |
|
MLB5 P |
2 |
|
|||||
|
MLA3 P |
0.537 |
|
CM1 G |
1.875 |
|
|||||
|
RD7 G |
0.509 |
|
RD7 A |
1.738 |
|
|||||
|
ALEPH 3 P |
0.476 |
|
ML7 A |
1.735 |
|
|||||
|
ML7 G |
0.441 |
|
RD 7 P |
1.724 |
|
|||||
|
CTS 2 P |
0.375 |
|
ALEPH 1 A |
1.720 |
|
|||||
|
continua en |
||||||||||
|
la página siguiente |
||||||||||
|
RD "A" P |
0.337 |
|
MLA3 A |
1.717 |
|
|||||
|
ALEPH 2 A |
0.290 |
|
CTS 2 A |
1.689 |
|
|||||
|
ML5 G |
0.248 |
|
RD7 G |
1.589 |
|
|||||
|
ALEPH 1 P |
0.200 |
|
ALEPH 2 A |
1.543 |
|
|||||
|
ML7 P |
0.063 |
|
ALEPH 3 A |
1.453 |
|
|||||
|
MLB5 P |
0.059 |
|
CM1 A |
1.438 |
|
|||||
|
Tabla 2a. Ranking de r² NISP/ MNE |
Tabla 2b. Ranking de la relación NSP/MNE |
|||||||||
Con el fin de observar en que medida los especímenes indeterminados a nivel elemento pero identificables a nivel guanaco, pinnípedos o aves oscurecían estas relaciones, calculamos la relación entre NSP [4] y MNE para cada uno de los sub-conjuntos -ver ranking en Tabla 2b. Esto nos permitirá observar el estado general de fragmentación de cada uno de los sub-conjuntos, considerando la cantidad total de elementos del mismo. En esta relación, cuanto mayor es el resultado, más incidencia del total de especímenes recuperados posee la muestra.
En la Figura 2 relacionamos los valores de r² NISP/MNE y de NSP/MNE para cada uno de los conjuntos. Puede observarse que parece no existir una tendencia general en las muestras, aún cuando es posible pensar que existen diferencias en el estado de fragmentación en cada una. Con el fin de explorar esto, definimos arbitrariamente tres grupos -Tabla 4- de acuerdo con tres rangos de valores tanto para r² NISP/MNE como para la relación NSP/MNE.
|
Q de muestras |
Rango de r² NISP/MNE |
|
| 14 |
1 a .8000 |
|
| 6 |
.7999 a .6000 |
|
| 6 |
.5999 a .4000 |
|
| 5 |
.3999 a .2000 |
|
| 2 |
.1999 a 0 |
|
| Q total = 33 |
||
Aquellos sub-conjuntos con altos valores de r² NISP/MNE y bajos valores de NSP/MNE presentarían baja fragmentación con una alta determinación de elementos. Opuestamente, los que presentan valores bajos en el primer caso y altos para el segundo, mostrarían alta fragmentación y poca determinación de especímenes.
.
| r² NISP/MNE |
Relación NSP/MNE |
Q de muestras |
Sub-Conjunto |
| 1 a .7000 |
1 a 2,499 |
10 |
RD"A" (aves). ML5 (aves), MLA3 (aves), Aleph 1 (aves), Aleph 3 (aves), CM1 (guanaco y aves), CTS 2 (aves), RD7 (pinnípedos y aves). |
| .6999 a .4000 | 2.5 a 4.999 |
15 |
CTS2 (guanaco), Aleph 1 (guanaco), Aleph 2 (guanaco y pinnípedos), Aleph 3 (guanaco y pinnípedos), RD"A" (guanaco), RD7 (guanaco), MLA3 (pinnípedos), MLB5 (guanaco y aves), ML5 (pinnípedos), ML7 (guanaco y aves), CM1 (pinnípedos). |
| .3999 a 0 |
> 5 |
8 |
Aleph 1 (pinnípedos), Aleph 2 (aves), ML5 (guanaco), MLB5 (pinnípedos), ML7 (pinnípedos), MLA3 (guanaco). CTS2 (pinnípedos), RD"A" (pinnípedos). |
De esto surgen una serie de datos interesantes. De los 10 sub-conjuntos arqueo-faunísticos que forman el primer grupo de la Tabla 4, 8 corresponden a aves (Q total = 11). En el segundo grupo, el guanaco está presente en 8, en tanto que en el tercero, 5 son de pinnípedos. De acuerdo con esto podemos decir que la tendencia general de los restos óseos de aves en el norte de Península Mitre presenta un menor grado de fragmentación que los demás subconjuntos. Por su parte, guanaco y pinnípedos presentan un mayor grado de fragmentación -levemente superior en el último caso [5] .
| |
|
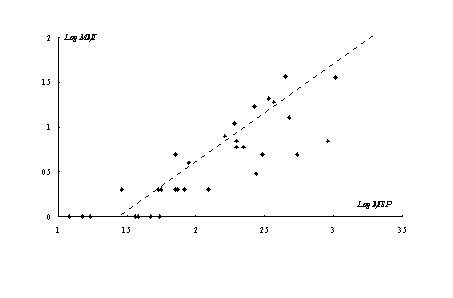
En la Figura 3 graficamos la relación entre NISP y MNI [6] de todos los sub-conjuntos analizados para el norte de Península Mitre (r = 0.695, r² = 0.483, p = .00001)(ver discusión en Grayson 1984:49-85). Puede observarse que la tendencia general de la región es la esperada, aunque levemente influenciada por los valores del NISP.
Huellas, Marcas y Restos Oseos
El estudio de huellas y marcas en restos óseos es una vía importante para explorar el grado de integridad de las muestras arqueológicas (Brain 1981 , Binford 1981 , Lyman 1994 ). En este estudio estamos interesados en conocer como se conjugan las relaciones entre marcas naturales y huellas culturales [7] en los diferentes sub-conjuntos arqueofaunísticos del norte de Península Mitre -en un sentido amplio. Nos interesa reconocer, a un nivel regional, cual es la incidencia de los procesos naturales y culturales en los restos óseos y si existe o no una representación diferencial de huellas y marcas en los distintos sub-conjuntos [8] .
De acuerdo con lo arriba expuesto, presentamos en primer término las relaciones entre Marcas y NSP, Huellas y NSP, y Huellas/Marcas -Tabla 5 y Apéndice, Tablas A7 - 9. A valores mayores, mayor es la presencia de marcas y/o huellas en cada uno de los sub-conjuntos. En el caso de la relación Huellas/Marcas, los valores más altos indican mayor presencia de Huellas en el sub-conjunto [9] .
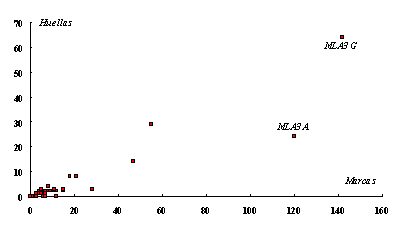
En Figura 4 puede observarse la relación Marcas y Huellas en los diferentes sub-conjuntos analizados. Se observa que MLA3 G y MLA3 A se separan del resto de las muestras. Esto puede ser un problema que tenga que ver con el tamaño de las mismas -ver el caso en Figura 1.
|
|
Figura 4. |
En la Figura 5 confrontamos la relación Huellas/Marcas con Marcas/NSP. Pretendemos observar posibles distorsiones que las frecuencias de marcas naturales mostraron cada vez que las huellas no estaban presentes en un sub-conjunto. Parecerían darse tres grupos distintos. Uno formado por aquellos sub-conjuntos en los que las marcas naturales dominan -por ausencia de huellas. Otro que presenta relaciones medias entre ambos índices y finalmente un tercero -más vago- con relaciones altas. La Tabla 6 presenta el detalle de los grupos mencionados.
|
Sub Conjunto |
Relación Marcas Naturales/NSP [10] |
RM |
Relación Huellas Culturales/NSP |
RH |
Relación Huellas/Marcas |
RG |
||
|
CM1 G |
0.333 |
1 |
0.133 |
3 |
0.400 |
7 |
||
|
ALEPH 2 G |
0.294 |
2 |
0.176 |
2 |
0.600 |
1 |
||
|
ALEPH 1 G |
0.250 |
3 |
0.000 |
25 |
0.000 |
25 |
||
|
CTS2 P |
0.241 |
4 |
0.034 |
12 |
0.143 |
22 |
||
|
ALEPH 3 G |
0.235 |
5 |
0.058 |
7 |
0.250 |
13 |
||
|
ALEPH 3 P |
0.220 |
6 |
0.097 |
4 |
0.444 |
6 |
||
|
CTS2 G |
0.188 |
7 |
0.063 |
6 |
0.333 |
9 |
||
|
MLB5 A |
0.173 |
8 |
0.018 |
17 |
0.107 |
24 |
||
|
MLA3 A |
0.157 |
9 |
0.023 |
15 |
0.200 |
17 |
||
|
RD"A" G |
0.145 |
10 |
0.036 |
10 |
0.250 |
14 |
||
|
ML5 A |
0.139 |
11 |
0.041 |
9 |
0.298 |
10 |
||
|
RD7 G |
0.135 |
12 |
0.036 |
10 |
0.167 |
21 |
||
|
RD"A" P |
0.111 |
13 |
0.027 |
14 |
0.250 |
15 |
||
|
MLA3 G |
0.101 |
14 |
0.071 |
5 |
0.451 |
5 |
||
|
MLB5 P |
0.089 |
15 |
0.200 |
1 |
0.200 |
17 |
||
|
CM1 A |
0.068 |
16 |
0.009 |
22 |
0.133 |
23 |
||
|
ML7 G |
0.064 |
17 |
0.000 |
25 |
0.000 |
25 |
||
|
RD"A" A |
0.061 |
18 |
0.000 |
25 |
0.000 |
25 |
||
|
ML7 A |
0.057 |
19 |
0.022 |
16 |
0.381 |
8 |
||
|
ML7 P |
0.057 |
20 |
0.000 |
25 |
0.000 |
25 |
||
|
MLB5 G |
0.056 |
21 |
0.016 |
18 |
0.286 |
11 |
||
|
ALEPH 1 P |
0.056 |
22 |
0.028 |
13 |
0.500 |
3 |
||
|
ALEPH 1 A |
0.047 |
23 |
0.010 |
21 |
0.222 |
16 |
||
|
ALEPH 3 A |
0.041 |
24 |
0.011 |
20 |
0.273 |
12 |
||
|
RD7 A |
0.034 |
25 |
0.007 |
23 |
0.200 |
19 |
||
|
CTS2 A |
0.031 |
26 |
0.000 |
25 |
0.000 |
25 |
||
|
MLA3 P |
0.031 |
27 |
0.053 |
8 |
0.527 |
2 |
||
|
ML5 P |
0.029 |
28 |
0.015 |
19 |
0.500 |
4 |
||
|
RD7 P |
0.023 |
29 |
0.004 |
24 |
0.182 |
20 |
||
|
ML5 G |
0.023 |
30 |
0.000 |
25 |
0.000 |
25 |
||
|
CM1 P |
0.014 |
31 |
0.000 |
25 |
0.000 |
25 |
||
|
ALEPH 2 P |
0.000 |
32 |
0.000 |
25 |
0.000 |
25 |
||
|
ALEPH 2 A |
0.000 |
32 |
0.000 |
25 |
0.000 |
25 |
Tabla 5. Ranking de las relaciones Marcas Naturales/NSP (RM), Huellas Culturales/NSP (RH) y Huellas/Marcas (RG) en los sub-conjuntos arqueofaunísticos analizados. Ver Apéndice, Tablas A7 - 9.
Figura 5. Relación Marcas/NSP vs. Huellas/Marcas. Datos procedentes de Tabla 5
De lo expuesto podemos resumir que las muestras arqueofaunísticas del norte de Península Mitre parecen mostrar una tendencia hacia una mayor presencia general de marcas que de huellas. Puede observarse que son pocos -8/33- los sub-conjuntos en los cuales las huellas culturales dominan proporcionalmente. No existe ningún sub-conjunto de aves, en tanto que guanaco y pinnípedos lo están con el mismo número de muestras.
| Q de Muestras |
Relación Huellas/Marcas vs. Marcas/NSP |
Sub-Conjunto |
| 8 |
alta |
MLA3 (pinnípedos y guanaco), ML5 (pinnípedos), Aleph 1 (pinnípedos y guanaco), Aleph 3 (pinnípedos), Aleph 2 (guanaco), CM1 (guanaco). |
| 16 |
media |
MLB5 (pinnípedos, guanaco y aves), Aleph 1 (aves), Aleph 3 (guanaco y aves), RD7 (pinnípedos, guanaco y aves), CTS2 (pinnípedos y guanaco), ML5 (aves), RDA (pinnípedos y guanaco), MLA3 (aves), CM1 (aves) |
| 9 |
baja |
Aleph 2 (pinnípedo y aves), CM1 (pinnípedos), ML5 (guanaco), CTS2 (aves). ML7 (pinnípedos, guanaco y aves), RDA (aves). |
Binford (1981:217 y ss.) ha utilizado la relación entre la frecuencia de epífisis proximales y distales de húmero y tibia de diferentes mamíferos terrestres para evaluar el grado de destrucción por carnívoros de conjuntos faunísticos. En las Figuras 6 (desthum.cht) y 7 (desttib.cht) mostramos estas relaciones para el caso de guanaco. Puede observarse que la mayoría de las muestras poseen en ambos casos, niveles confiables de no destrucción. Se destacan -en el caso del húmero- Aleph 2 -único dentro del área de destrucción- y ML7. En el caso de la tibia, CTS2 y CM1 son los que se encuentran fuera del área de no destrucción. Esto mostraría, que en líneas generales, las muestras de guanaco presentan un bajo nivel de destrucción por carnívoros [11] .
Figura 6. Relación entre frecuencia de epífisis proximales y distales de húmero de guanaco en el norte de Península Mitre.
Figura 7. Relación entre frecuencia de epífisis proximales y distales de tibia
de guanaco en el norte de Península Mitre.
Densidad Osea e Integridad
Los estudios de densidad ósea (Lyman 1984, 1991 y 1994 , Lyman et al. 1992 , Elkin y Zanchetta 1991 , Kreutzer 1992 , Chambers 1992 -en Lyman 1994-) han demostrado ser una vía de análisis importante para conocer el grado de integridad del registro faunístico. Discutiremos aquí los sub-conjuntos faunísticos tomando los datos de densidad ósea realizados por Elkin (Elkin y Zanchetta 1991, Elkin 1990 ) para guanaco y aves. Para el caso de los pinnípedos emplearemos los valores de Chambers (1992 en Lyman 1994) [12] .
En la Tabla 7 presentamos las correlaciones entre %MAU y densidad ósea para los sub-conjuntos de guanaco y pinnípedos. Arbitariamente hemos calificado a aquellas muestras con valores de r² .2500 como de baja correlación, entre .2501 y .5000 como media en tanto que aquellas con valores .5001 como de alta correlación.
Con el fin de ejemplificar algunos de los casos mencionados en la tabla precedente -ya que hacerlo con cada uno de ellos sería sumamente tedioso- hemos realizado algunas figuras en donde mostramos diferentes casos. Las primeras dos figuras -8 y 9- muestran casos en donde la correlación entre %MAU y la densidad de los restos óseos es baja.
|
Sub-Conjunto |
r² |
p |
Correlación con densidad ósea |
|
|
Bahía |
Thetis |
|||
|
ALEPH 1 G |
.2825 |
.6432 |
media |
|
|
ALEPH 1 P |
.5529 |
.1497 |
alta |
|
|
ALEPH 2 G |
.3074 |
.6259 |
media |
|
|
ALEPH 2 P |
.5485 |
.4691 |
alta |
|
|
ALEPH 3 G |
.0716 |
.7323 |
baja |
|
|
ALEPH 3 P |
.0013 |
.9159 |
baja |
|
|
CTS 2 G |
.2500 |
.6667 |
baja |
|
|
CTS 2 P |
.0926 |
.6957 |
baja |
|
|
Ea. María |
Luisa |
|||
|
ML5 G |
.2578 |
.0765 |
media |
|
|
ML5 P |
.0101 |
.7116 |
baja |
|
|
ML7 G |
.0021 |
.9004 |
baja |
|
|
ML7 P |
.0667 |
.6749 |
baja |
|
|
MLA3 G |
.0054 |
.7799 |
baja |
|
|
MLA3 P |
.0410 |
.4572 |
baja |
|
|
MLB5 G |
.0106 |
.7379 |
baja |
|
|
MLB5 P |
.0697 |
.4070 |
baja |
|
|
continua en la |
||||
|
página siguiente |
||||
|
Rancho |
Donata |
|||
|
CM1 G |
.2168 |
.3524 |
baja |
|
|
CM1 P |
.0698 |
.5271 |
baja |
|
|
RD "A" G |
.1735 |
.3047 |
baja |
|
|
RD "A" P |
.4159 |
.5538 |
media |
|
|
RD7 G |
.0094 |
.7634 |
baja |
|
|
RD7 P |
.0067 |
.7710 |
baja |
|
Tabla 7. Correlaciones entre % MAU y densidad ósea para los sub-conjuntos de guanaco (G) y pinnípedos (P)
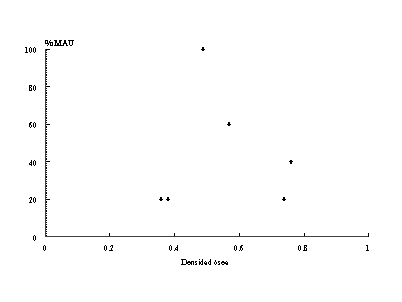
Figura 8. Gráfico de dispersión entre %MAU y densidad ósea para el sub-conjunto
de pinnípedos en Aleph 3.
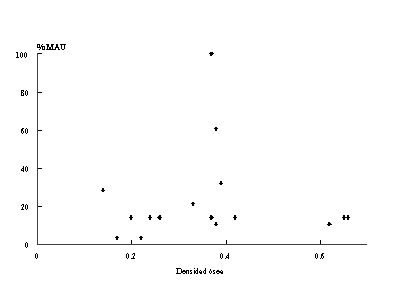
Figura 9. Gráfico de dispersión entre %MAU y densidad ósea para el sub-conjunto
de guanaco en MLA 3.
Por su parte, la tercera -Figura 10- muestra una correlación media correspondiente a la muestra de guanaco de ML5. El caso de las aves ha sido tratado recientemente por Savanti (1994). Ella ha discutido los diferentes casos aquí tratados (ver Savanti 1994:63-97). Ella concluye que la conservación diferencial de partes esqueletarias de las aves tiene estrecha relación con la densidad ósea global de los elementos recuperados. Aquí, y a modo de ejemplo mostramos el caso de CM1 -Figura 11. Estos estudios realizados por Savanti se basaron en los valores de densidad de Elkin (1990 ), calculados sobre un ejemplar de Chloephaga melanoptera. Al carecer de valores específicos para cada una de las especies de nuestra región, partimos del supuesto que las relaciones entre los valores de las densidades de cada elemento óseo son proporcionales en las diferentes especies avifaunísticos [13] .
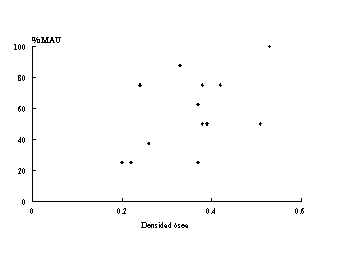
Figura 10. Gráfico de dispersión entre %MAU y densidad ósea para el sub-conjunto de guanaco en ML5.
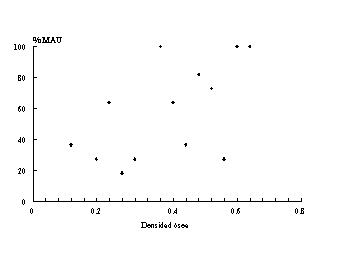
Figura 11 Gráfico de dispersión entre %MAU y densidad ósea para el sub-conjunto
de aves en CM1.
Basándonos en Savanti (1994) reagrupamos los distintos sub-conjuntos avifaunísticos según su dependencia con la densidad ósea -Tabla 8.
|
Sub-Conjunto |
Dependencia con |
|
Aves |
densidad ósea |
|
ALEPH 1 |
alta |
|
ALEPH 2 |
alta |
|
ALEPH 3 |
alta |
|
CTS 2 |
alta |
|
MLA3 |
alta |
|
MLB5 |
media |
|
ML5 |
media |
|
ML7 |
media |
|
CM1 |
media |
|
RD "A" |
media |
|
RD7 |
alta |
Tabla 8. Tipo de correlación de los distintos sub-conjuntos avifaunísticos en el norte de Península Mitre. Basado en Savanti 1994.
Puede observase que, en general, todos los sub-conjuntos avifaunísticos presentan una alta dependencia con la densidad. La única localidad en la cual todas las muestras poseen una tendencia igual entre ellas es Bahía Thetis.
Calibrando la Integridad de las Muestras
En los apartados anteriores, presentamos una serie de análisis tendientes a comprender la integridad de las muestras óseas en el norte de Península Mitre. Cada uno de ellos presenta distintas características de las propiedades de los materiales faunísticos. Sintéticamente, podemos resumirlas de la siguiente manera:
la tendencia a la fragmentación de los restos óseos parece ser diferencial entre aves, guanacos y pinnípedos. Las primeras muestran un bajo grado general de fragmentación general.
la evidencia de procesos naturales en los restos óseos parece ser más importante que la de los procesos culturales.
desde el punto de vista de la densidad ósea, 6 de las 11 muestras de aves y una de pinnípedos se correlacionan con la misma, en tanto que el guanaco no posee correlaciones altas.
baja intensidad de la acción de carnívoros sobre restos de guanacos y quizás también sobre pinnípedos.
Estas características son las que, de alguna manera nos alertan sobre propiedades generales del registro faunístico de la región, e incluso entendemos que podrían extenderse a otras de la Isla Grande de Tierra del Fuego. Sin embargo, existen peculiaridades que deben aclararse con el fin de calibrar adecuadamente las mismas.
El punto que consideramos importante es ponderar apropiadamente las muestras. Con esto queremos decir que, tras la amplia y diversa gama de procesos naturales y culturales involucrados en la formación del registro, no podemos asumir que todos las muestras arqueológicas poseen el mismo peso para entender estrategias y tácticas en poblaciones humanas. Muy por el contrario, creemos que es más adecuado pensar que las mismas tienen un valor diagnóstico diferencial. Por lo tanto es necesario calibrar en que medida nos informan tanto de procesos naturales como culturales. Con el fin de tratar de acercarnos a esto, hemos resumido en la Tabla 9, algunas de las propiedades de las muestras arqueofaunísticas discutidas anteriormente.
La variables que hemos tomado son A) la relación entre r²NISP/MNE vs NSP/MNE; B) el grado de dependencia en la densidad ósea; C) la relación Huellas/Marcas vs. Marcas/NISP; y D) el ranking general de la relación entre Huellas/Marcas. La tabla mencionada debe leerse de la siguiente manera: para A) poca, media y mucha indican la fragmentación de la muestra; para B) alto, medio y bajo señalan a correlación con la densidad ósea; y para C) alta, media y baja indican la mayor incidencia de marcas naturales sobre huellas culturales y D) rankea los sub-conjuntos de acuerdo con la mayor presencia general de huellas sobre las marcas.
En la Tabla 9 hemos agrupado los sub-conjuntos por localidad arqueológica. Puede observarse que Rancho Donata es la que presenta la menor cantidad de muestras con ranking alto de RG -salvo CM1 G [14] . Ea. María Luisa y Bahía Thetis poseen tanto valores altos como bajos en ese ranking. Esto podría indicar tentativamente que los materiales recuperados en ambientes boscosos -ya sea el bosque joven o el parque fueguino- poseen una mayor presencia de huellas culturales.
|
r²NISP/MNE vs |
Correlación con |
Huellas/Marcas vs |
||
|
Sub-Conjunto |
NSP/MNE* |
densidad ósea |
Marcas/NSP+ |
RG# |
|
A |
B |
C |
D |
|
|
Bahía Thetis |
||||
|
ALEPH 1 A |
poca |
alta |
media |
|
|
ALEPH 1 G |
media |
media |
alta |
|
|
ALEPH 1 P |
mucha |
baja |
alta |
|
|
ALEPH 2 A |
mucha |
alta |
baja |
|
|
ALEPH 2 G |
media |
baja |
alta |
|
|
ALEPH 2 P |
media |
media |
baja |
|
|
ALEPH 3 A |
poca |
alta |
media |
|
|
ALEPH 3 G |
media |
baja |
media |
|
|
ALEPH 3 P |
media |
alta |
alta |
|
|
CTS 2 A |
poca |
alta |
baja |
|
|
CTS 2 G |
media |
media |
media |
|
|
CTS 2 P |
mucha |
media |
media |
|
|
Ea. María Luisa |
||||
|
ML5 A |
poca |
media |
media |
|
|
ML5 P |
media |
alta |
alta |
|
|
ML5 G |
mucha |
media |
baja |
|
|
ML7 A |
media |
media |
baja |
|
|
ML7 P |
mucha |
media |
baja |
|
|
ML7 G |
media |
media |
baja |
|
|
MLA3 A |
poca |
alta |
media |
|
|
MLA3 G |
mucha |
media |
alta |
|
|
MLA3 P |
media |
media |
alta |
|
|
|
página siguiente |
|||
|
MLB5 A |
media |
media |
media |
|
|
MLB5 G |
media |
media |
media |
|
|
MLB5 P |
mucha |
alta |
media |
|
|
Rancho Donata |
||||
|
CM1 A |
poca |
media |
media |
|
|
CM1 G |
poca |
baja |
alta |
|
|
CM1 P |
media |
alta |
baja |
|
|
RD "A" G |
media |
baja |
media |
|
|
RD "A" P |
mucha |
baja |
media |
|
|
RD "A" A |
poca |
media |
baja |
|
|
RD7 A |
poca |
alta |
media |
|
|
RD7 G |
media |
alta |
media |
|
|
RD7 P |
poca |
alta |
media |
|
Antes de continuar, queremos aclarar que no existe una solución ideal en el estudio de la integridad. Muy por el contrario, la simple lectura de la tabla anterior nos alerta sobre la multiplicidad de situaciones ante las cuales nos enfrentamos. No discutiremos aquí cada una de las muestras [15] , por el contrario, estamos tratando de observar si diferentes propiedades del registro arqueofaunístico, y algunas de sus relaciones, nos permiten separar muestras con grados distintos de integridad. Para ello rankeamos las combinaciones posibles de las tres propiedades de Tabla 10 [16] -los diferentes rankings se encuentran en el Apéndice, Tablas A10 -15.
De alguna manera, la combinación ideal de un sub-conjunto es:
| A= |
poca | B= |
bajo | C= |
alta | RG= |
alto |
Esto indicaría que el sub-conjunto se encuentra poco fragmentado, la supervivencia de elementos parece no estar relacionada con la densidad ósea y con poca incidencia de las marcas naturales. Cuanto más cercano a 1 es la posición en RG, una mayor posibilidad de estudiar actividad humana estaría evidenciada en la muestra. No existe ningún sub-conjunto arquefaunístico que cumpla con estas cuatro posibilidades dentro de los estudiados.
En la Tabla 10 resumimos los rankings resultantes de las seis posibles combinaciones postuladas para cada uno de los sub-conjuntos -ver detalle en Apéndice Tablas A10 - A15.
Se pueden observar algunas peculiaridades. Dos sub-conjuntos, CM1 G y Aleph 2 A siempre se ubican en las mismas posiciones en los distintos rankings. En Aleph 2 A su baja posición indica que
es alta la fragmentación
la incidencia de las marcas naturales es alta
los especímenes recuperados se correlacionan con la densidad ósea
existe una mayor evidencia de marcas que de huellas culturales.
Esto señala que Aleph 2 A sería una muestra adecuada para discutir distintos procesos de formación natural. Por el contrario, CM1 G parecería ser un buen sub-conjunto para efectuar análisis de tácticas y estrategias. En el resto de las muestras pueden observarse otras particularidades. Hay algunos sub-conjuntos que se mantienen agrupados, cualquiera sea el criterio que tomemos como constante al efectuar el ranking. Los mismos son:
|
i) |
ML5 A y CM1 A |
|
ii) |
Aleph 1 A y Aleph 3 A |
|
iii) |
Aleph 3 G y RD"A" G |
|
iv) |
Aleph 1 G y MLA3 P |
|
v) |
CTS 2 G, MLB5 A y MLB5 G |
|
vi) |
Aleph 2 P, ML7 A y ML7 G |
|
vii) |
Aleph 3 P y ML5 P |
|
viii) |
ML7 P y ML5 G |
|
Ranking |
Ranking |
Ranking |
Ranking |
Ranking |
Ranking |
|
|
Sub-Conjunto |
A,B,C,D |
A,C,B,D |
B,A,C,D |
B,C,A,D |
C,A,B,D |
C,B,A,D |
|
Tabla A10 |
Tabla A11 |
Tabla A12 |
Tabla A13 |
Tabla A14 |
Tabla A15 |
|
|
CM1 G |
1 |
1 |
1 |
1 |
1 |
1 |
|
ML5 A |
2 |
2 |
7 |
10 |
9 |
12 |
|
CM1 A |
3 |
3 |
8 |
11 |
10 |
13 |
|
RD "A" A |
4 |
9 |
9 |
16 |
25 |
25 |
|
ALEPH 1 A |
5 |
4 |
23 |
24 |
11 |
18 |
|
ALEPH 3 A |
6 |
5 |
24 |
25 |
12 |
19 |
|
MLA3 A |
7 |
6 |
24 |
26 |
13 |
20 |
|
RD7 A |
8 |
7 |
25 |
27 |
16 |
21 |
|
RD7 P |
9 |
8 |
26 |
28 |
15 |
22 |
|
CTS 2 A |
10 |
10 |
27 |
31 |
26 |
31 |
|
ALEPH 2 G |
11 |
11 |
2 |
2 |
2 |
2 |
|
ALEPH 3 G |
12 |
16 |
3 |
4 |
16 |
9 |
|
RD "A" G |
13 |
17 |
4 |
5 |
17 |
10 |
|
ALEPH 1 G |
14 |
12 |
10 |
7 |
3 |
4 |
|
MLA3 P |
15 |
13 |
11 |
8 |
4 |
5 |
|
CTS 2 G |
16 |
18 |
12 |
12 |
18 |
14 |
|
MLB5 A |
17 |
19 |
13 |
13 |
19 |
15 |
|
MLB5 G |
18 |
20 |
14 |
14 |
20 |
16 |
|
ALEPH 2 P |
19 |
22 |
15 |
17 |
27 |
26 |
|
ML7 A |
20 |
23 |
16 |
18 |
28 |
27 |
|
ML7 G |
21 |
24 |
17 |
19 |
29 |
28 |
|
ALEPH 3 P |
22 |
14 |
28 |
22 |
5 |
7 |
|
ML5 P |
23 |
15 |
29 |
23 |
6 |
8 |
|
RD7 G |
24 |
21 |
30 |
29 |
21 |
23 |
|
CM1 P |
25 |
25 |
31 |
32 |
30 |
32 |
|
ALEPH 1 P |
26 |
26 |
5 |
3 |
7 |
3 |
|
RD "A" P |
27 |
28 |
6 |
6 |
22 |
11 |
|
MLA3 G |
28 |
27 |
18 |
9 |
8 |
6 |
|
CTS 2 P |
29 |
29 |
14 |
15 |
23 |
17 |
|
ML7 P |
30 |
31 |
20 |
21 |
31 |
29 |
|
ML5 G |
30 |
31 |
20 |
20 |
31 |
29 |
|
MLB5 P |
32 |
30 |
32 |
30 |
24 |
24 |
|
ALEPH 2 A |
33 |
33 |
33 |
33 |
33 |
33 |
Tabla 10. Rankings de las combinaciones de las propiedades de los sub-conjuntos arqueofaunísticos.
Cada uno de estos grupos posee distintas posiciones en los rankings y por lo tanto son relevantes para diferentes discusiones arqueológicas. El caso i) indica la presencia de una baja fragmentación con dependencias medias en la densidad de los huesos y en las relaciones con marcas y huellas. El caso levemente opuesto a éste parece ser el del grupo iv) en dónde la fragmentación de las muestras es mayor que en i). En ii) es interesante destacar que si bien la fragmentación de los sub-conjuntos es baja, el grado de dependencia en la densidad de los huesos es alta. Las muestras de iii) -ambas de guanaco- dependen muy poco de la densidad ósea, y sus valores para las otras propiedades es medio. El grupo v) presenta valores medios en general, en tanto que vi) solo en el caso de la densidad, siendo bajos en las otras propiedades. Un grupo interesante es el vii) -ambos de pinnípedos- ya que al rankear tomando como constante las variables relacionadas con huellas y marcas, los sub-conjuntos se ubican en posiciones altas. En los otros rankings, la posición desciende. Finalmente, el grupo viii) posee niveles bajos en todos los casos.
Además de estos casos de agrupamientos de los sub-conjuntos, existen otras particularidades. Hay sub-conjuntos ubicados en posiciones altas cuando tomamos las columnas B o C -ver Tabla 9.- pero al tomar la fragmentación como primera propiedad del ranking, los encontramos en posiciones medias o bajas -i.e., Aleph 2 G, Aleph 1 P, RD"A" P MLA3 G. Esto nos alerta muy claramente sobre la relevancia diferencial de las muestras para discutir determinados aspectos.
Considerando en que medida los diferentes sub-conjuntos se ubican en determinadas posiciones podemos agruparlos de acuerdo con sus posibles grados de integridad semejante [17] . Los mismos serían:
| Grados de Integridad | Sub-conjunto |
| Ideal |
no hay |
| Superior |
CM1 G |
| Mayor |
ML5 A, CM1 A, Aleph 2 G, Aleph 1 G, MLA3 P |
| Medio |
RD"A" A, Aleph 1 A, Aleph 3 A, CTS2 G, MLB5 A, MLB5 G, Aleph 3 P, ML5 P, Aleph 1P, RD"A" P, MLA3 G, RD"A" G, Aleph 3 G |
| Bajo |
MLA3 A, RD7 A, RD7 P, CTS2 A, Aleph 2 P, ML7 A, ML7 G, RD7 G, CM1 P, CTS2 P, ML7 P, ML5 G, MLB5 P |
| Muy Bajo |
Aleph 2 A |
| Integridad | Aves | Pinnípedos | Guanaco | Bosque joven | Parque | Pradera |
|
Superior |
0 |
0 |
1 (9)[3] |
1 (7)[3] |
0 |
0 |
|
Mayor |
2 (19)[6] |
1 (9)[3] |
2 (18)[6] |
3 (21)[9] |
2 (17)[6] |
0 |
|
Medio |
4 (36)[12] |
4 (36)[12] |
5 (45)[15] |
6 (40)[18] |
4 (33)[12] |
3 (50)[9] |
|
Bajo |
4 (36)[12] |
6 (55)[18] |
3 (27)[9] |
4 (27)[12] |
6 (50)[18] |
3 (50)[9] |
|
Muy Bajo |
1 (9)[3] |
0 |
0 |
1 (7)[3] |
0 |
0 |
Tabla 12. Distribución por grados de integridad de los sub-conjuntos de acuerdo con tipo de animales y ambientes. Los valores entre paréntesis indican porcentajes por columna. Los valores entre corchetes indican porcentajes de acuerdo al total de la muestra (Q=33).
En la Tabla 12 hemos agrupado los diferentes sub-conjuntos de acuerdo con los distintos tipos de fauna y ambientes.
En líneas generales, y considerando grados de integridad superior y mayor, podemos decir que los sub-conjuntos faunísticos de guanaco son los más representados -9% del total-, seguidos por las aves -6%. Teniendo en cuenta grados de integridad bajos y muy bajos, los sub-conjuntos de pinnípedos son los más representados (18%), en tanto que las aves suman el 15%. Desde el punto de vista de los ambientes, los boscosos jóvenes tienden a presentar los mejores grados de integridad (12%) [18] .
Para finalizar, queremos decir que debemos ser claros y entender las distintas escalas de análisis implicadas en el estudio de los procesos de formación [19] . Al tener nuestro estudio una escala regional, nos interesa conocer las tendencias en esa escala. Una vez entendida ésta, la escala micro -sitio- cobra una relevancia mayor, ya que puede ser calibrada en función de expectativas determinadas, y no tan solo mediante la simple identificación de determinados procesos y/o agentes.
Agradecimientos
Al Museo del Fin del Mundo, Ushuaia, por el apoyo brindado al proyecto mencionado. A Luis A. Borrero, A. Sebastián Muñoz , Juan B. Belardi y Daniel Olivera por sus comentarios.
Bibliografía
Binford, Lewis R.
1981 Bones. Ancient Men and Modern Myths. Academic Press, New York.
1984 Faunal Remains from Klasies River Mouth. Academic Press. Orlando.
Bondel, Conrado Santiago
1988 Geografía de Tierra del Fuego. Guía docente para su enseñanza. Museo Territorial. Ushuaia.
Borrero, Luis A.
1988 Estudios tafonómicos en Tierra del Fuego: Su relevancia para entender procesos de formación del registro arqueológico. En Arqueología Contemporánea Argentina H. Yacobaccio ed. pp 13-32. Ed. Búsqueda. Buenos Aires.
Brain, C.K.
1981 The Hunters or the Hunted? The University of Chicago Press. Chicago
Elkin, Dolores C.
1990 Informe Final de Beca de Iniciación. CONICET. MS.
Elkin, Dolores C. & J.R. Zanchetta
1991 Densitometría Osea de Camélidos. Aplicaciones arqueológicas. Shincal 3(1): 195-204.
Grayson,Donald K.
1978 On the Quantification of vertebrate archaeofaunas. Advances in Archaeological Method and Theory 2:199-237.
1984 Quantitative Zooarchaeology: Topics in the Analysis of Archaeological Faunas. Academic Press. New York.
1991 Alpine faunas from the White-Mountains, California: Adaptative change in the Late Prehistoric Great Basin? Journal of Archaeological Science 18:483-506.
Klein, Richard & K. Cruz-Uribe
1984 The Analysis of Animals Bones from Archaeological Sites. University of Chicago Press. Chicago.
Kreutzer, Lee Ann
1992 Bison and deer bone mineral densities: comparisons and implications for the interpretation of archaeological faunas. Journal of Archaeological Science 19:271-294
Lanata, José Luis
1990a Human, Terrestrial and Sea Mammals at Peninsula Mitre, Tierra del Fuego. En Hunters of the Recent Past L. Davis & B.O.K. Reeves eds., pp. 400-406. Unwin Hyman. London.
1990b Discussing Maritime Adaptation at Southeastern Tierra del Fuego. En Abstracts Sixth International Conference. E.A. Myler, B.D. Smith & M.A. Zeder eds., pg..86. International Council for Archaeozoology, Washington D.C.
1993 Estados Alterados. Procesos de formación y conjuntos faunísticos en Rancho Donata, Tierra del Fuego. En Explotación de Recursos Faunísticos en Sistemas Adaptativos Americanos. J.L. Lanata comp., pp. 163-176. Arqueología Contemporánea 4. Buneos Aires.
1996 Propiedades en la depositación de valvas de moluscos en muestras arqueológicas del sudeste fueguino. Trabajo presentado en Fourh International Congress of Medical and Applied Malacology - Santiago Chile.
Lanata, José Luis, Marcelo Weissel, Soledad Caracotche, Juan B. Belardi, Sebastián A. Muñoz y Florencia Savanti.
1992 20000 Huesos de viaje submarino: análisis faunístico del sitio Bahía Crossley. Isla de los Estados. Palimpsesto 1: 9-24.
Lanata, José Luis & Alejandro Winograd
1988 Gritos y Susurros: Aborígenes y Lobos Marinos en el litoral de la Tierra del Fuego. En Arqueología de las Américas. 45° Congreso Internacional de Americanistas. pp. 227-246. Bogotá.
Lefèvre, Christine
1989 L'Avifaune de Patagonie Australe et ses Relations avec l'Homme au Cours des Six Derniers Millenaires. Tesis doctoral. Université de Paris. Paris I. Paris.
Lyman, R. Lee
1984 Bone density and differential survivorship in fossil classes. Journal of Antropological Archaeology 3:259-299.
1991 Prehistory of the Oregon Coast. The Effects of Excavations Strategies and Assemblage Size on Archaeological Inquiry. Academic Press. San Diego.
1994 Vertebrate Taphonomy. Cambridge University Press. Cambridge.
Lyman, R. Lee, Lori E. Houghton & Annell L. Chambers
1992 The effect of structural density on marmot skeletal part representation in archaeological sites. Journal of Archaeological Science 19:557-573.
Mengoni G., Guillermo L.
1988 Análisis de materiales faunísticos de sitios arqueológicos. Xama 1: 71-120.
Muñoz, A. Sebastián
1994 Arqueofaunas de la Costa Atlántica Fueguina. Marcas Naturales y Huellas Culturales. Tesis de Licenciatura. Facultad de Filosofía y Letras. Universidad de Buenos Aires. Buenos Aires.
Muñoz, A. Sebastián y Florencia Savanti
1994 Observaciones tafonómicas sobre restos avifaunísticos de la costa noreste de Tierra del Fuego. Presentado al XI Congreso Nacional de Arqueología Argentina. San Rafael. Mendoza
Richardson, P.
1980 Carnivore damage to antelope bones and its archaeological implications. Paleontologia Africana 23:109-125.
Savanti, Florencia
1994 Las Aves en la Dieta de los Cazadores-Recolectores Terrestres de la Costa Fueguina. Temas de Arqueología. PREP-CONICET. Buenos Aires.
Silveira, M. & M. Fernández
1988 Huellas y marcas en el material óseo de Fortín Necochea (Partido de Gral. La Madrid, Prov. de Bs. Aires). En De Procesos, Contextos y Otros Huesos. N. Ratto & A. Haber eds., pp. 45-52 Facultad de Filosofía y Letras. UBA. Buenos Aires.
NOTAS
[1] Distintos estudios se han realizado sobre la fauna de los diferentes sitios de la región (i.e., Lanata 1990a, 1993, 1996, Lanata y Winograd 1988, Savanti 1994, Muñoz 1994, Borella nd, y existen otros en preparación), en los cuales el análisis fue a nivel específico. Remitimos al lector a los mismos para un mayor detalle. En el caso de las aves agrupamos a todos los géneros y especies recuperados -sin embargo ver estudios específicos en los trabajos citados. Ello se debe a que nos interesa conocer como se comportan los huesos de las aves desde el punto de vista de las propiedades del registro arqueológico tratadas en este trabajo. Por otra parte, nuestro interés aquí se centra en conocer tendencias regionales en la integridad de los registros faunísticos, para lo cual las categorias analíticas empleadas -más amplias- son pertinentes. Aclaramos también que no utilizaremos, en este estudio, los estadios de meteorización de los restos óseos. Esto se basa en que tan solo tenemos muestras actualísticas para guanaco (Borrero 1988). Estudios actualísticos sobre aves y mamíferos marinos se encuentran en marcha (Muñoz y Savanti 1994, Borella nd), por lo que serán incorporados en el futuro.
[2] Destacamos dos sub-conjuntos en la Figura 1, MLA3 G (guanaco) y MLA3 A (aves). Ambos se encuentran dentro de la tendencia general de la región, tan solo que son las muestras más grandes de la misma.
[3] Existe una diferencia en el cálculo de esta relación en los trabajos mencionados. Richardson (1980) calcula la relación NISP/MNE dividida x 100. Los otros autores directamente correlacionan la cantidad de especímenes reconocidos para cada elemento con el número mínimo de elementos necesarios para dar cuenta de la muestra considerando tanto especímenes enteros como fragmentados (Binford 1984).
[4] Utilizamos la terminología de Grayson (1991:487, NSP= total number of specimens). En este caso, incluímos aquellos especímenes que no pudieron ser adscriptos a un elementos determinado pero si fue posible adjudicarlos a alguno de los sub-conjuntos.
[5] La costilla constituye el elemento que mayor fragmentación presenta en las diferentes muestras.. ACA PONER ALGO EN EL FINAL DE LAS CONCLUSIONES PUES ESTO CONTRADICE LO QUE ESPERAMOS PARA GUANACO Y LOBO
[6] El MNI se calculó considerando los estados de fusión ósea (no fusionados y fusionados).
[7] Seguimos aquí la terminología utilizada por Silveira y Fernández (1988).
[8] En este trabajo no discutiremos tácticas de procesamiento de las distintas faunas, ya que no es nuestro objetivo específico. Para este tipo de análisis ver Lanata 1988, 1990b, 1993, Lanata et al. 1992, Lanata y Winograd 1988, Savanti 1994, Muñoz 1994.
[9] Esta relación tan solo toma en cuenta la dicotomía natural/cultural. Se agrupan en estas categorías las diferentes huellas y marcas registradas en las muestras (ver Apéndice).
[10] Empleamos el NSP ya que se identificaron huellas y marcas en especímenes que no pudieron ser adjudicados a elementos determinados, pero si a las categorias de guanaco, pinnípedos y aves aquí utilizadas.
[11] Esto es aplicable para carnívoros del tamaño de un Canis o mayor. Esto incluye a los Pseudalopex sp. que podemos encontrar en la región.
[12] En otros trabajos se han presentado detalles de la aplicación de la densidad ósea de sub-conjuntos particulares (ver Savanti 1994, Muñoz 1994), y otros se encuentran en curso.
[13] Savanti (1994:95) nos alerta sobre la posible existencia de diferencias entre las distintas especies en relación con los valores de la densidad ósea. El caso más significativo puede estar dado por los pinguinos, los que podrían tener valores de densidad ósea muy distintos al resto de las aves fueguinas -aunque cormoranes y petreles pueden constituírse en otros ejemplos. Como en nuestro caso nos interesa conocer la tendencia general de las aves, optamos por considerar a todas las especies avifaunísticas como un recurso, tomando para ello los datos de densidad ósea de Elkin (1990) realizados sobre un ejemplar de Chloephaga melanoptera. Cuando contemos con nuevos valores para otras especies procederemos a atomizar más adecuadamentes nuestros estudios de densidad ósea sobre las aves de la región.
[14] Uno de los factores a tener en cuenta en Rancho Donata, es el estado de meteorización. Ya hemos mencionado que aquí no abordaremos este estudio -ver nota 1. Sin embargo, podemos adelantar que esta localidad sería la que presenta un mayor grado de meteorización general de los restos óseos. Esto puede deberse al alto contenido de arena -silicatos- en los sedimentos, lo que provocaría un aceleramiento en el proceso de destrucción del hueso.
[15] Como ya hemos mencionado anteriormente, análisis particulares se han realizado y/o están siendo llevado a cabo.
[16] Combinamos estas tres propiedades en distintas formas ya que el orden de cada uno de los rankings resultantes se modifica de acuerdo a cual propiedad se toma como constante. El ranking de la relación Huellas/Marcas (RG) se lo utilizará para separar -cuando sea necesario- dos o más casos con iguales combinaciones de A, B y C. Si bien es esperable que estos rankings den resultados diferentes, es también posible que aquellos en los cuales una misma propiedad -A, B o C- se mantenga constante, presente ordenes relativamente semejantes.
[17] Este primer intento de evaluación de la integridad de las muestras considera los grados para la región en estudio exclusivamente.
[18] Ya hemos mencionado anteriormente, que nuestras muestras de bosque -tanto joven como del parque fueguino- son más en número que las de pradera. Sin embargo, esta tendencia sería la esperable.
[19] Luego de haber pasado por una farragosa y tediosa descripción de valores, relaciones, tablas y figuras, creemos que es necesario aclarar algunos puntos sobre los restos de avifauna.Hemos puntualizado que la tendencia de las aves en la región es presentar una baja fragmentación (ver también Lanata 1990b, 1993, Lanata et al. 1992, Savanti 1994, Lefèvre 1989) con una alta dependencia en la densidad ósea. En la Tabla 9 puede observarse que de las 10 primeras posiciones en los rankings que toman a la fragmentación como primer ordenador, 8 son sub-conjuntos de aves. Además, 5 de ellos presentan mucha dependencia en la densidad ósea -posiciones entre 23 y 31. De alguna manera, la poca fragmentación así como la dependencia con la densidad y una baja presencia de huellas son esperables por las características de las aves. En otros trabajos (Lanata et al. 1992, Lanata 1993, ver también Lefèvre 1989 y Savanti 1994) discutimos el procesamiento de éstas, por lo que no nos reiteraremos aquí. El punto que queremos destacar es que, en la medida que los restos de aves cumplan con estas tendencias, se hace difícil separar factores naturales de culturales que han contribuido en su formación. Estudios tafonómicos -i.e., Muñoz y Savanti 1994 - nos ayudarán en el futuro a ajustar estos problemas en las muestras avifaunísticas.