 Especial NAyA Volumen 2
Especial NAyA Volumen 2
ANÁLISIS FAUNÍSTICO DE LOS RESTOS EXCAVADOS EN LAS CASAS DE TERRIN (PANAMÁ LA VIEJA): UNA APROXIMACIÓN A LA DIETA Y A LA ECOLOGÍA*
Richard Cooke***
RESUMEN
Este documento ofrece avances en el análisis de los restos de fauna recuperados durante la excavación de un pozo colonial en la antigua ciudad de Panamá La Vieja. Ofrece una aproximación descriptiva y ecológica: de llevar cada hueso al nivel taxonómico más preciso y - especialmente en lo que a peces se refiere - procurar inferir dónde y, con menos certeza, cómo fueron capturados. Esta información permite acercarse a los patrones alimenticios de los habitantes de esta ciudsad (en cuanto a selectividad y acceso a recursos se refiere) y establecer comparaciones con los contextos arqueológicos prehispánicos de la región.
INTRODUCCIÓN
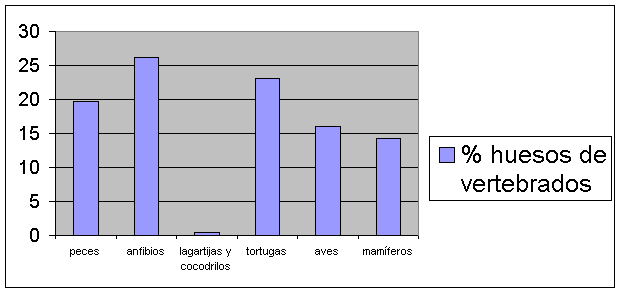
Las excavaciones en el pozo colonial[2] aportaron una muestra grande de huesos de vertebrados los cuales están siendo analizados por los autores en el Laboratorio de Arqueozoología del Instituto Smithsonian de Investigaciones Tropicales, localizado en la calzada de Amador. Una buena parte de los huesos pertenecen a tres especies domésticas de mamíferos, que arribaron a Panamá con los españoles en el siglo XVI de nuestra era: el caballo (Equus caballus), la vaca (Bos taurus) y el puerco (Sus scrofa). Aún no hemos terminado su análisis. En este documento presentamos un informe preliminar sobre otros vertebrados que ya hemos identificado taxonómica y anatómicamente: peces (264 huesos), anfibios (351 huesos), tortugas de agua dulce (aproximadamente 310 huesos), aves (216 huesos) y mamíferos (192 huesos) (Cuadros 1 y 2). Exceptuando la muestra de peces, la diversidad taxonómica es baja: una sola especie de anfibio (el ubicuo sapo común [Bufo marinus]); abundantes pedazos de caparazones de jicotea (Trachemys scripta), una tortuga de agua dulce; gallinas traídas del Viejo Mundo y patos que parecen ser de origen americano (Cairina); perros y gatos domésticos; dos abundantes roedores que llegaron a América en pos del ser humano: el ratón casero (Mus musculus) y la rata negra (Rattus rattus); y, por último, unas cuantas especies de la fauna silvestre, como la iguana verde (Iguana iguana), el venado de cola blanca (Odocoileus virginianus) y el saíno (Tayassu tajacu), las cuales eran populares fuentes de carne en tiempos prehispánicos (Cooke y Ranere 1992, Cooke et al. 1985) y deberían haber sido abundantes en las sabanas, bosques de galería y rastrojos en las afueras de Panamá La Vieja durante toda la época colonial cuando la población humana era considerablemente menor que en tiempos prehispánicos.
En vista de que faltan por analizar los restos de los animales que indiscutiblemente proporcionaban la mayor cantidad de carne a Panamá La Vieja, nuestra primera preocupación ha sido descriptiva y ecológica: llevar cada hueso al nivel taxonómico más preciso y - especialmente en lo que a peces se refiere - procurar inferir dónde y, con menos certeza, cómo fueron capturados. Los índices de abundancia que hemos utilizado son el número de especímenes identificados (NEI) y el número mínimo de individuos (NMI). Cuando este último se desprende del elemento anatómico más abundante - independientemente de los tamaños de los individuos representados - se le denomina NMI-1. Cuando está supeditado a la atribución de los huesos de un taxón a rangos de tamaños, se le llama NMI-2[3]. No hemos recurrido a ecuaciones alométricas para estimar los pesos de los individuos aunque dicho método puede ser más preciso que el método ocular comparativo con tal de que los huesos arqueológicos estén completos y bien preservados y de que se tenga a mano una muestra de esqueletos de individuos de diferentes edades, lo bastante grande como para asegurar buenos valores R2. Creemos que una investigación alométrica y morfométrica de la muestra de huesos de aves de corral y gatos domésticos sería de especial interés porque se tiene poca información sobre el tipo de gallinas y gatos que llegaron al trópico americano y sobre las características genéticas y morfométricas de los patos.
En los cuadros, los pesos que damos en gramos se derivan de la comparación visual de cada hueso con los de esqueletos de animales de los mismos taxones que fueron colectados con datos biométricos. El término biomasa es la suma de los pesos estimados de todos los individuos de cada taxón calculado con base en el NMI-1 o, si la muestra lo amerita, al NMI-2. Cuando se hayan analizado todos los huesos, se presentará información adicional sobre el aporte relativo de cada grupo de vertebrados a la dieta y vida cotidiana de la comunidad de Panamá La Vieja.
PECES Y PESCA
La muestra de peces sumó 264 huesos de los cuales 16 (6%) pertenecen a tiburones (Chondrichthyes, Elasmobranchii) y los demás 248 (94%) a peces teleósteos (Osteichthyes, Actinopterygii). Doscientos veinte huesos (86%) fueron identificados hasta familia, 214 (84%) hasta género y 189 (74%) hasta especie (Cuadros 1 y 3). Estos porcentajes de huesos taxonómicamente significativos son más altos que los que se han reportado para muestras de huesos de peces de sitios precolombinos en Panamá, situación que se desprende, a lo mejor, de las especiales condiciones tafonómicas del pozo colonial, p.ejm., el rápido cubrimiento de los restos al ser arrojados en el pozo seguido por bajos niveles de perturbación pos-deposicional.
Taxonomía
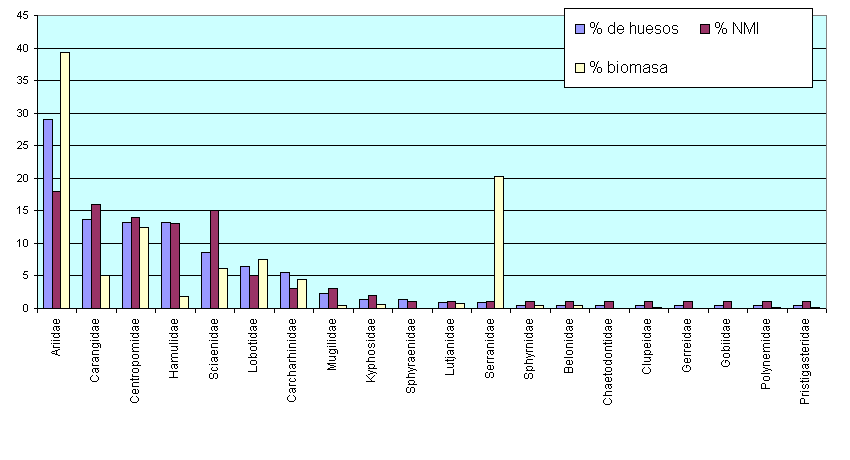
Están representados 20 familias, 40 géneros, 43 especies y seis especies identificadas tentativamente (estas últimas están señaladas en los cuadros por el prefijo cf). Todos estos taxones son de origen marino aunque algunas especies, p. ejm., el tiburón azulito (Carcharhinus leucas), un congo (Cathorops tuyra), los róbalos negro y blanco (Centropomus nigrescens y C. viridis) y la mojarra bandera (Eucinostomus currani) remontan ríos hasta agua completamente dulce donde pueden ser pescados con trasmallos y anzuelos (Cooke y Tapia 1994a; Vásquez y Thorson 1982). Las cinco familias mejor representadas aportaron conjuntamente el 75% de los huesos de peces: los bagres marinos (Ariidae) (29%), los jureles (Carangidae) (13,6%), los róbalos (Centropomidae) (13,2%), las corvinas (Sciaenidae) (13,2%) y los berrugates (Lobotidae) (6%) (Cuadros 4 y 5). Estas proporciones subrayan por sí mismas el enfoque litoral de la pesca. Las cuatro primeras familias contienen muchas especies por lo que no es de sorprenderse que se hayan identificado seis barbudos marinos (Ariidae), ocho jureles (Carangidae), seis roncadores (Haemulidae) y ocho corvinas y coscochas (Sciaenidae). Cinco taxones se reportan por primera vez en muestras arqueológicas de vertebrados en el Pacífico panameño: un pez mariposa (Chaetodontidae: Chaetodon humeralis) (Chaetodontidae), un roncador (Haemulidae: Anisotremus taeniatus), chopas (Kyphosidae: Kyphosus), la salema (Kyphosidae: Sectator ocyurus) y un gallote (Sciaenidae: Paralonchurus petersi). (Los nombres populares que usamos son los que se escuchan con mayor frecuencia en Panamá; ver, también, Cooke y Ranere 1994).
Ecología y zonas de pesca
En lo que al Pacífico tropical oriental se refiere, la mayor parte de las especies comprendidas en las 20 familias representadas se encuentran en las aguas poco profundas de estuarios, ensenadas, lagunas costeras o playas arenosas. Unas cuantas prefieren sustratos duros, como arrecifes y zonas rocosas, en tanto que otras evitan fondos suaves, aguas turbias con sedimentos suspendidos o corrientes de baja salinidad. Los juveniles, sub-adultos y adultos pequeños de algunos taxones, como los pargos, muestran preferencia por hábitats litorales y fluviales y se trasladan hacia aguas más profundas y claras a medida que crecen. Algunos depredadores de la fauna nectónica que alcanzan tamaños grandes, como los pargos, meros y longjinos o aguirres (Oligoplites altus), son bastante oportunistas e incursionan en muchos tipos de hábitats a fin de aprovechar la abundancia periódica de sus alimentos primarios.
Dadas estas consideraciones, se comprende por qué nuestra atribución de los taxones representados a cuatro zonas del ambiente marino no constituye una clasificación rígida, sino un esfuerzo por inferir en cuáles hábitats se hubieran dado las mayores probabilidades de capturar las especies de peces consumidas en Panamá La Vieja con técnicas pre-industriales (Cuadro 6). Hemos empleado la siguiente clave:
Negritas (p.ejm., Carcharhinus leucas) este es el hábitat donde existen las mayores probabilidades de capturar esta especie regularmente.
Letras normales (p.ejm., Selene oerstedii) esta especie es lo suficientemente abundante en este hábitat como para ser pescado allí ocasionalmente.
Itálica (p.ejm., Rhizoprionodon longurio) esta especie suele entrar en este biomo en etapas tempranas de su ciclo de vida.
Letras en mayúscula (p,.ejm., HAEMULOPSIS ELONGATUS) - esta especie suele ocupar este biomo en etapas tardías de su ciclo de vida[4].
Estuario bajo y medio
El estuario bajo y medio comprende las zonas mareales de ríos y quebradas, sus desembocaduras, los manglares litorales y la zona de mezcla de agua dulce y salada con fondos predominantemente fangosos (Day et al. 1989; Cooke 1992). Se estima que las 21 especies que abundan en esta zona proporcionaron el 53 % de los huesos de peces, el 51 % de las especies, el 46 % de los individuos y el 63 % de la biomasa del pescado. Muy significativo es el aporte de algunas especies que alcanzan un buen tamaño, como el bagre gris (Selenaspis dowi) y los robalos (Centropomus spp.). Según nuestros estimados, los ocho individuos de bagre gris en esta muestra sumaron un peso de 67 Kg (el 34% de la biomasa de peces). Los 14 róbalos identificados arrojaron una biomasa de 24.7 Kg, lo que significa un peso promedio de 1.76 Kg por individuo. También eran consumidos en Panamá La Vieja berrugates (Lobotes surinamensis) de 1.5 a 5.3 Kg y corvinas boquiamarillas (Cynoscion albus) de hasta 4.5 Kg El individuo de mayor peso (40 Kg), no obstante, es un mero (Epinephelus) que no pudo ser identificado hasta especie. Aunque la pesca de meros y cabrillas grandes se practica hoy en día en aguas de más de 10 metros de profundidad Cooke y Tapia (1994b) reportaron un espécimen de Epinephelus analogus de 20 Kg en el atajo del estero Palo Blanco en Aguadulce por lo que es posible que el único mero cuyos restos se depositaron en el pozo colonial haya sido pescado en aguas someras estuarinas.
Hoy en día, estas y otras especies que habitan el estuario bajo y medio son capturadas con trasmallos y anzuelos, métodos que habrían sido eficaces en los alrededores de Panamá La Vieja.
Estuario alto
Según nuestras investigaciones en la Bahía de Parita, algunas especies que se encuentran en estuarios del Pacífico tropical oriental evitan las zonas más turbias y someras siendo pescadas, más bien, en el borde externo de la zona de mezcla de aguas donde el estuario sensu stricto se topa con el mar abierto (Cooke 1992). Nos cuentan los pescadores de los pueblos costeros de Boca de Parita y El Rompío, que el roncador Pomadays panamensis se obtiene hoy en día en agua de 5 a 15 brazas, zona que es preferida, también, por el pargo de la mancha (Lutjanus guttatus), la corvina pelona (Cynoscion squamipinnis) y por dos barbudos marinos (Arius troschelii [5] y A. dasycephalus). Aunque datos anecdóticos como estos no garantizan que los individuos de estas especies y de las otras que destacamos en la columna correspondiente del Cuadro 6 hayan sido capturados definitivamente en esta zona, su presencia sugiere que los pescadores que abastecían de pescado a Panamá La Vieja llevaban cayucos hacia lo más profundo del estuario para pescar, tal vez con anzuelos.
Playas arenosas y corrientes de agua clara
La segunda especie en importancia en Panamá La Vieja, de acuerdo al NEI y al NMI-2 es el ñao-ñao (Orthopristis chalceus). Este pequeño roncador, que raras veces sobrepasa los 500 g, se congrega en cardúmenes muy grandes. En algunos sitios precolombinos, como Sitio Sierra, era consumido con frecuencia (Cooke y Ranere 1994, 1999). Aunque es una especie de aguas litorales, evita aguas turbias, fondos fangosos y, probablemente, la baja salinidad (Allen y Robertson 1994: 150; Cooke y Tapia 1994b), al igual que la cojinúa (Caranx caballus), catarnica (Selene peruviana), barracuda o picuda (Sphyraena ensis) y aguja (Tylosurus cocodrilus). Un estudio de una trampa de barrera erigida en una playa arenoso-cascajoso en Veracruz, provincia de Panamá, en la década de 1970 (DCroz et al. 1977) demostró que estos aparejos son aptos para pescar cojinúas, sierras (Scomberomorus sierra) y catarnicas por lo que es imprudente inferir que la presencia de estas especies y las otras que agrupamos en la columna correspondiente del Cuadro 6 alude necesariamente al uso de embarcaciones, señuelos y redes.
Rocas y arrecifes
De los taxones de peces reportados en esta muestra los que menos propensión muestran por aguas litorales son la boquipenda (Caranx speciosus), las chopas (Kyphosus spp.), la salema (Sectator ocyurus) y el pez mariposa (Chaetodon humeralis). De estos, el último pertenece a una familia cuyas especies están asociadas por lo general a arrecifes coralinos. Las boquipendas se alimentan cerca del fondo (Allen y Robertson 1994: 127). Al igual que las chopas y salemas son abundantes cerca de islas. Cada uno de estos taxones está representado en la muestra por solo un individuo. A lo mejor, se pescaron en la cercanía de las pequeñas islas, como Naos y Perico, donde los galeones atracaban en tiempos coloniales.
REPTILES
Se encontraron cuatro taxones de reptiles en el pozo. El más abundante, con creces, es la jicotea (Emydidae: Trachemys scripta), especie de tortuga dulceacuícola que es en extremo frecuente en Panamá donde quiera que haya agua dulce de cierta profundidad. Doscientos fragmentos pudieron ser identificados anatómicamente; se hallaron, además, aproximadamente 110 pedazos diversos de caparazón y plastron.
En tiempos precolombinos, la jicotea era consumida regularmente en sitios aledaños a la Bahía de Parita (Cooke y Ranere 1989, 1992) y, en la actualidad, es aún perseguida por su carne. De los 200 elementos anatómicamente significativos hallados en el pozo, se identificaron tan sólo un hueso craneal y 14 pos-craneales. De estos, nueve pertenecieron al mismo individuo de aproximadamente 600 g, que bien pudo haber encontrado una muerte natural en el pozo. Si excluimos este individuo, sin embargo, el dominio de huesos del caparazón sobre los del resto del cuerpo hace pensar que lo que se tiraba en el pozo era este elemento con uno que otro hueso adherido. Es frecuente encontrar jicoteas muertas durante el verano cuando se secan las charcas y ciénagas; sin embargo, creemos que si esta muestra de huesos hubiera resultado de la recolección y descarte de animales recogidos en el monte, o si todos los animales hubiesen muerto por causas naturales en el pozo, se hubieran encontrado muchísimo más huesos craneales y pos-craneales. Aunque no se observaron evidencias del descuartizamiento, como cortaduras y ralladuras, si se hierve una jicotea entera el caparazón se desbarata facilitando la utilización de la carne. El elemento anatómico más abundante en la muestra (el hioplastrón izquierdo) arroja un NMI-1 de 15 individuos. Se estima en base a comparaciones del tamaño de los plastrones arqueológicos con los de esqueletos de jicoteas en la muestra de referencia del STRI, que los 15 individuos se agrupan en la siguiente manera:
| Peso aproximado (g): |
3000 |
2500 |
2200 |
2000 |
1500 |
| Individuos: |
1 |
4 |
2 |
5 |
3 |
A estos se agregan tres individuos cuyos plastrones, al parecer, no sobrevivieron (de 1000 g, 600 g y 400 g respectivamente). Esto da un NMI-2 de 18 individuos y una biomasa para esta especie de aproximadamente 33.9 Kg Ya una proporción alta del peso de una tortuga está representada por el caparazón, se calcula que tan sólo el 40% es carne. En este caso, de haber sido consumidas como alimento humano, las jicoteas hubieran proporcionado aproximadamente 13.56 Kg de carne.
Otros reptiles
Se encontró un diente de alguna especie de cocodrilo (Cocodrylidae).
No poseemos un esqueleto del tamaño del individuo que representa por lo que
nuestro estimado de su peso (>15 Kg) es muy tentativo. Se hallaron tres
huesos de un sólo individuo, de aproximadamente 80 g, de la lagartija que
se conoce en Panamá como borriguero (Ameiva ameiva), especie comensal
y amante de áreas abiertas, cuyos tejidos, contrariamente a la creencia popular,
no son tóxicos y tienen buen sabor. Un individuo de la iguana verde (Iguana
iguana) que aportó tres huesos pesaba unos 2000g.
ANFIBIOS
La única especie de anfibio hallada en el pozo es el sapo común (Bufo marinus) que aportó 351 huesos pertenecientes a animales con pesos estimados entre los 50 y 240 g. El elemento más abundante (la tibio-fíbula derecha) arroja un NMI-1 de 27 individuos. Aunque, seguramente, el NMI-2 es bastante mayor - debido al rango de pesos estimados- hace falta una revisión más completa de la muestra para poder calcularlo con certeza.
Huesos del sapo común son frecuentes en arqueofaunas precolombinas de Coclé y Azuero. (Fernández de Oviedo constata que, por 1520, los indígenas de Nicaragua y Panamá consumían como alimento una especie cuya descripción coincide perfectamente con Bufo marinus [ver referencias incluidas en Cooke 1989]). En base a la proporcionalidad de grupos anatómicos de huesos de sapo hallados en Sitio Sierra (Coclé), en un basurero fechado en aproximadamente 100-300 d.C., Cooke (1989) infirió que esta especie era utilizada por los habitantes de este pueblo si bien no fue posible confirmar que se la comían (pudo haber sido usada para fines rituales). Es poco probable que los europeos residentes en Panamá La Vieja comieran Bufo marinus, una especie morfológicamente parecida al Bufo bufo de Europa que es rechazado enfáticamente como alimento humano. Sin embargo, en vista de que debería haber existido un remanente de población de origen indígena en Panamá La Vieja durante el periodo en el que se depositaron los huesos en el pozo, cabe en lo posible que el sapo común (cuyos tejidos musculares no son tóxicos [Cooke 1989]) hubiera servido de alimento humano.
El siguiente cuadro compara el número de huesos que pertenecen a cuatro zonas anatómicas del sapo común que, de acuerdo a experimentos realizados por Cooke (1989), son pertinentes a la preparación de los anfibios para ser comidos. Estas son, tal como se presenta a continuación (1) la cabeza, (2) los suspensorios y la columna vertebral, (3) las patas delanteras y (4) las patas traseras. (Ver, también, el Cuadro 7).
| Zona anatómica |
Hueso |
Núm de huesos por individuo |
Núm. de huesos generados por 29 individuos |
Núm. de huesos hallados en el pozo |
% de huesos hallados con referencia al número hipotético |
| Cabeza |
Dentario Escamoso Etmoides Frontal Maxila Nasal Parasfenoides Pterigoides |
2 2 1 2 2 2 1 2 |
58 58 29 58 58 58 29 58 |
18 1 10 1 2 2 1 16 |
|
| Total: |
12 |
406 |
51 |
12.6% |
|
| Suspensorios y Vértebras |
Coracoides Escápula Ilión Uróstilo Vértebras |
2 2 2 1 9 |
58 58 58 29 261 |
2 15 34 9 77 |
|
| Total: |
16 |
464 |
137 |
29.5% |
|
| Patas delanteras |
Húmero Radio-Cúbito |
2 2 |
58 58 |
29 28 |
|
| Total: |
4 |
116 |
57 |
49.1% |
|
| Patas traseras |
Fémur Tibio-fíbula |
2 2 |
58 58 |
36 42 |
|
| Total: |
4 |
116 |
78 |
67.2% |
Estos cuadros señalan que los huesos del cráneo están subrepresentados de manera significativa en tanto que la tasa de supervivencia de huesos de las partes más comestibles (las patas) sobrepasa la del basurero precolombino de Sitio Sierra (Cooke 1989: figura 3). En teoría, pues, la proporcionalidad de partes anatómicas supervivientes no se compagina con una situación de mortandad natural, p.ejm., si los sapos quedaban atrapados en el pozo y no podían salir. Proponemos, en conclusión, que los huesos del sapo común que se depositaron en el pozo colonial pudieron haber procedido de animales culturalmente modificados.
AVES
Se hallaron 216 huesos de aves. De estos 39 no proveyeron información taxonómica específica aunque una vértebra pertenece a alguna especie pequeña con un peso aproximado de 350 g. Es posible que represente una paloma (Columbidae). Diez huesos de patos (Anatidae) representan por lo menos dos individuos. Aunque la colección de referencia de esqueletos del STRI contiene bastantes especies de patos silvestres carece de formas domésticas lo que inhibe la precisión de este análisis taxonómico. Sin embargo, un húmero proximal es tan parecido a un espécimen del pato real (Cairina moschata) hallado en un basurero indígena en Sitio Sierra y fechado entre el 100 y 300 d.C., que suponemos que representa esta especie. El ornitólogo-paleontólogo Storrs Olson que examinó el hueso precolombino (Cooke 1984) supuso que pertenecía a un animal doméstico. Los demás huesos de pato en el pozo son bastante más robustos que los del pato doméstico común de Europa (Anas platyrhynchos) y exhiben algunas diferencias morfológicas con respeto al esqueleto de este.
Esperamos poder mejorar el nivel de análisis taxonómico y morfológico de los huesos de patos tan pronto hayamos conseguido una muestra representativa de esqueletos de formas domésticas de Cairina moschata y Anas platyrhynchos.
Ciento sesenta y cinco huesos son de gallina (Gallus). La muestra contiene animales de todas las edades y representa más de siete individuos. Este hecho en adición a la fecha del depósito resalta la importancia de un análisis arqueofaunístico más minucioso en el futuro.
Un húmero fue asignado a otra especie doméstica: el pavo (Meleagris gallopavo). Es probable que un cúbito también sea de pavo. Esta especie fue domesticada originalmente por los indígenas precolombinos de Mesoamérica. Hasta la fecha no se ha reportado en sitios panameños aunque es bastante frecuente en la Península de Nicoya (Kerbis 1973), región que fue asentada varios siglos antes del contacto español por gentes mexicanas.
Un húmero distal exhibe características parecidas a un espécimen de la pava silvestre (Cracidae: Penelope purpurascens) aunque es más grande. Sospechamos que pertenece al pavón (Cracidae: Crax rubra), especie que está ausente en nuestra colección de referencia de esqueletos. Su presencia, de ser comprobada, indicaría que la gente que proveía de carne a Panamá La Vieja cazaba en bosques maduros.
MAMÍFEROS
Mencionamos en nuestra introducción que está por terminar el complejo y demorado análisis de la muestra de huesos de vacas, caballos y puercos sin cuyos resultados no podemos inferior el aporte de cada grupo de vertebrados a la vida cotidiana y dieta de los habitantes de Panamá La Vieja.
Los 192 huesos de mamíferos que no representan estas especies pertenecen a seis taxones. Dos de ellos, el venado de cola blanca (Odocoileus virginianus) y el saíno (Tayassu tajacu), son silvestres. Los 24 huesos de venado pertenecen a por lo menos cuatro individuos, dos adultos y dos inmaduros. Los cuatro elementos de saíno son de un solo individuo adulto. Se hallaron nueve huesos de roedores de la familia americana Cricetidae.
Dos especies domésticas están representadas: el perro y el gato, siendo el segundo mucho más abundante. Dos elementos de perro pertenecen a un individuo adulto y otro inmaduro. Cuarenta y ocho elementos de gato representan por lo menos cinco individuos, dos adultos y tres inmaduros. Un análisis detallado de la secuencia de fusión de las epífisis aumentaría, seguramente, el número de individuos.
Como es de esperarse en un ambiente urbano, se hallaron bastantes huesos de dos roedores del Viejo Mundo que llegaron a América en los barcos europeos: el ratón doméstico (Mus musculus) y la rata negra (Rattus rattus). A fin de identificar estos especímenes pedimos prestado un esqueleto de cada una de estas especies al Instituto Smithsonian de Washington y, también, un espécimen de la segunda rata comensal, Rattus norvegicus. Por lógica, se requeriría una muestra más representativa de esqueletos de todas las edades para poder separar las dos especies de ratas en todos los casos. Sin embargo, nos parece probable que la muestra de huesos hallados en el pozo represente únicamente la variedad negra, que es menos robusta que la rata de Noruega. Los cien huesos de esta especie representan por lo menos nueve individuos (NMI-1) y los seis de ratón, un solo individuo.
CONCLUSIÓN
Si bien este análisis ha abordado sólo una parte de la importante muestra de huesos de vertebrados hallados en el pozo colonial de Panamá La Vieja, nos ofrece un panorama interesante del aprovechamiento y de la comensalidad de las especies de animales.
Al igual que durante el periodo precolombino, los pescadores que abastecían a la ciudad de pescado operaban principalmente en zonas litorales, como estuarios, bocanas, manglares y playas arenosas y pescaban especies que eran importantes en la época prehispánica y siguen siéndolo hoy en día, como bagres, congos, róbalos, roncadores, berrugates y corvinas. Es muy difícil inferir los métodos de pesca sin tener a mano evidencia sobre los tipos de artefactos que se habrían utilizado: investigaciones nuestras y de DCroz et al. sobre la pesca artesanal actual han señalado que algunos métodos de pesca bastante sencillos, como las trampas de barrera, son capaces de capturar la mayor parte de las especies identificadas en esta muestra, aún las más pesadas, como los bagres, róbalos, berrugates y meros. Sin embargo, el hecho de que los habitantes de Panamá La Vieja hayan consumido especies como el pargo blanco (Pomadasys panamensis), las chopas y salemas (Kyphosidae), las boquipendas y un pez mariposa aluden al uso de pequeñas embarcaciones a cierta distancia de la costa y a la pesca cerca de islas rocosas.
La presencia de venados de cola blanca, saínos e iguanas en la muestra indica que otro patrón precolombino continuó después del contacto con los españoles. Aunque la primera especie hubiese sido perseguida intensivamente por los cazadores prehispánicos, logró mantener sus niveles de población (Cooke 1992b; Cooke y Ranere 1989, 1992) por lo que se supone que era abundante en áreas cercanas a la ciudad colonial.
La muestra del pozo resalta la importancia en la dieta local de las aves de corral: gallinas y patos. En vista de que los huesos de estos taxones están en muy buen estado, son apropiados para un análisis más minucioso cuyo objetivo sería determinar el tamaño y edades de los individuos representados y, de ser posible, algunas características genéticas.
El hallazgo de más de 300 huesos de jicoteas y sapos no es de sorprenderse en un pozo de agua dulce y sería lógico presumir que sus esqueletos representan animales que murieron allí atrapados. Aún así, hemos señalado que la distribución de las partes anatómicas de estos taxones no es cónsona con un patrón de mortandad natural. Ambas especies eran alimentos populares en la época precolombina y sería imprudente descartar la posibilidad de que hubiesen sido consumidas por los sectores no-europeos de esta comunidad.
Por último, los abundantes huesos de ratas y ratones y de los gatos domésticos que los cazaban no representa sorpresa alguna en un ambiente urbano.
Cuadro 1
| Taxon |
Nombre común |
Núm. Familias |
Núm. Familias tentativas |
Núm. Géneros |
Núm. Especies |
Núm, especies tentativas |
Núm huesos |
% huesos |
Peso huesos (g) |
NMI-1 |
NMI-2 |
Biomasa estimada (g) |
| Chondrichthyes |
tiburones y rayas |
2 |
0 |
3 |
2 |
1 |
16 |
1,2 |
2,9 |
4 |
9300 |
|
| Osteichthyes |
peces óseos |
18 |
0 |
36 |
41 |
6 |
248 |
18,5 |
234 |
96 |
189.105 |
|
| Pises |
peces (total) |
20 |
0 |
39 |
43 |
7 |
264 |
19.7 |
236.9 |
100 |
198.405 |
|
| Anura |
ranas y sapos |
1 |
0 |
1 |
1 |
0 |
351 |
26,2 |
9,08 |
27 |
? |
? |
| Squamata |
lajartijas y cocodrilos |
3 |
0 |
3 |
3 |
0 |
7 |
0,5 |
8,83 |
3 |
3 |
? |
| Testudines |
tortugas |
1 |
0 |
1 |
1 |
0 |
310 |
23,1 |
1925,7 |
15 |
19 |
33,9 |
| Reptilia |
reptiles (total) |
4 |
0 |
4 |
4 |
0 |
317 |
23,7 |
1934,53 |
45 |
? |
? |
| Aves |
aves |
3 |
1 |
3 |
3 |
0 |
216 |
16,1 |
279,86 |
11 |
? |
? |
| Mammalia |
mamíferos |
6 |
0 |
6 |
6 |
0 |
192 |
14,3 |
539,3 |
22 |
? |
? |
| Totales: |
Total: |
34 |
1 |
53 |
57 |
7 |
1340 |
2999,67 |
Cuadro 2
Cuadro 3
| Contexto |
Clase/sub-clase |
cf |
Familia |
cf |
Género |
cf |
Especie |
Parte anatómica |
Posición de vértebra |
Izquierda |
Derecha |
Quemado |
No quemado |
NII |
Peso de los huesos(g) |
Peso estimado de los individuos (g) |
NMI |
Biomasa por taxón |
| pIII-2 |
Elasmobranchii |
Carcharhinidae |
vértebra |
0 |
0 |
0 |
2 |
2 |
0,3 |
0 |
||||||||
| pIII-1 |
Elasmobranchii |
Carcharhinidae |
vértebra |
0 |
0 |
0 |
1 |
1 |
0,1 |
0 |
||||||||
| A22 CII AT2 |
Elasmobranchii |
Carcharhinidae |
Carcharhinus |
leucas |
vértebra |
0 |
0 |
0 |
1 |
1 |
0,3 |
5000 |
1 |
5000 |
||||
| pIII-2 |
Elasmobranchii |
Carcharhinidae |
Rhizoprionodon |
longurio |
vértebra |
0 |
0 |
0 |
3 |
3 |
1,9 |
3000 |
1 |
3000 |
||||
| pIII-2 |
Elasmobranchii |
Carcharhinidae |
Rhizoprionodon |
longurio |
vértebra |
0 |
0 |
0 |
5 |
5 |
0,2 |
600 |
1 |
600 |
||||
| pIII-2 |
Elasmobranchii |
Sphyrnidae |
Sphyrna |
cf |
lewini |
vértebra |
0 |
0 |
0 |
4 |
4 |
0,1 |
700 |
1 |
700 |
|||
| pII-2 |
Actinopterygii |
fragmento |
0 |
0 |
0 |
2 |
2 |
0,7 |
||||||||||
| pII-4 |
Actinopterygii |
fragmento |
0 |
0 |
0 |
6 |
6 |
1,1 |
||||||||||
| pIII-1 |
Actinopterygii |
fragmento |
0 |
0 |
0 |
26 |
26 |
0,6 |
||||||||||
| pIII-2 |
Actinopterygii |
Ariidae |
vértebra |
0 |
0 |
0 |
1 |
1 |
0,09 |
|||||||||
| pIII-2 |
Actinopterygii |
Ariidae |
fragmento |
0 |
0 |
0 |
2 |
2 |
1,4 |
|||||||||
| pIII-2 |
Actinopterygii |
Ariidae |
"Arius"1 |
dasycephalus |
post-temporal |
0 |
1 |
0 |
1 |
1 |
0,1 |
250 |
1 |
250 |
||||
| pIII-2 |
Actinopterygii |
Ariidae |
"Arius" |
troschelii2 |
cleitron |
0 |
1 |
0 |
1 |
1 |
2,5 |
2000 |
||||||
| pIII-2 |
Actinopterygii |
Ariidae |
"Arius" |
troschelii |
epihial |
1 |
0 |
0 |
1 |
1 |
0,3 |
2000 |
||||||
| pII-2 |
Actinopterygii |
Ariidae |
"Arius" |
troschelii |
preopérculo |
0 |
1 |
0 |
1 |
1 |
0,2 |
2000 |
1 |
2000 |
||||
| pIII-2 |
Actinopterygii |
Ariidae |
"Arius" |
troschelii |
coracoides |
0 |
1 |
0 |
1 |
1 |
1,4 |
1300 |
||||||
| pIII-3 |
Actinopterygii |
Ariidae |
"Arius" |
troschelii |
epiótico |
1 |
0 |
0 |
1 |
1 |
1,1 |
1300 |
||||||
| pIII-2 |
Actinopterygii |
Ariidae |
"Arius" |
troschelii |
urohial |
0 |
0 |
0 |
1 |
1 |
0,4 |
1300 |
||||||
| pIII-2 |
Actinopterygii |
Ariidae |
"Arius" |
troschelii |
queratohial |
1 |
0 |
0 |
1 |
1 |
1,2 |
1300 |
1 |
1300 |
||||
| A22 CIII L |
Actinopterygii |
Ariidae |
"Arius" |
troschelii |
frontal |
1 |
0 |
0 |
1 |
1 |
0,3 |
450 |
1 |
450 |
||||
| pIII-2 |
Actinopterygii |
Ariidae |
"Arius" |
troschelii |
epiótico |
1 |
0 |
0 |
1 |
1 |
0,3 |
200 |
1 |
200 |
||||
| A22 CII AT2 pII-8 |
Actinopterygii |
Ariidae |
Bagre |
pinnimaculatus |
vértebra |
0 |
0 |
0 |
1 |
1 |
2,3 |
3500 |
1 |
3500 |
||||
| A22 CII AT2 |
Actinopterygii |
Ariidae |
Bagre |
pinnimaculatus |
frontal |
0 |
1 |
0 |
1 |
1 |
4,8 |
2300 |
1 |
2300 |
||||
| pIII-2 |
Actinopterygii |
Ariidae |
Cathorops |
espina dorsal o pectoral |
0 |
0 |
0 |
1 |
1 |
0,3 |
||||||||
| pIII-2 |
Actinopterygii |
Ariidae |
Cathorops |
cf |
hypophthalmus |
cleitron |
1 |
0 |
0 |
1 |
1 |
0,5 |
200 |
1 |
200 |
|||
| pIII-2 |
Actinopterygii |
Ariidae |
Cathorops |
multiradiatus |
cleitron |
1 |
0 |
0 |
1 |
1 |
0,1 |
30 |
1 |
30 |
||||
| pIII-2 |
Actinopterygii |
Ariidae |
Cathorops |
tuyra |
pteriogóforo dorsal, I |
0 |
0 |
0 |
1 |
1 |
0,1 |
300 |
1 |
300 |
||||
| pII-4 |
Actinopterygii |
Ariidae |
Selenaspsis |
dowii3 |
cuadrado |
0 |
1 |
0 |
1 |
1 |
2,1 |
13000 |
||||||
| pII-3 |
Actinopterygii |
Ariidae |
Selenaspsis |
dowii |
cuadrado |
1 |
0 |
0 |
1 |
1 |
1,9 |
13000 |
||||||
| pII-4 |
Actinopterygii |
Ariidae |
Selenaspsis |
dowii |
epihial |
2 |
0 |
0 |
2 |
2 |
5,2 |
13000 |
2 |
26000 |
||||
| pII-4 |
Actinopterygii |
Ariidae |
Selenaspsis |
dowii |
epiótico |
0 |
1 |
0 |
1 |
1 |
2,8 |
13000 |
||||||
| pII-4 |
Actinopterygii |
Ariidae |
Selenaspsis |
dowii |
esfenótico |
1 |
0 |
0 |
1 |
1 |
1,4 |
13000 |
||||||
| pII-4 |
Actinopterygii |
Ariidae |
Selenaspsis |
dowii |
hiomandíbula |
0 |
1 |
0 |
1 |
1 |
1,6 |
13000 |
||||||
| pII-4 |
Actinopterygii |
Ariidae |
Selenaspsis |
dowii |
hiomandíbula |
1 |
0 |
0 |
1 |
1 |
1,4 |
13000 |
||||||
| pII-4 |
Actinopterygii |
Ariidae |
Selenaspsis |
dowii |
post-temporal |
0 |
1 |
0 |
1 |
1 |
2,6 |
13000 |
||||||
| pII-4 |
Actinopterygii |
Ariidae |
Selenaspsis |
dowii |
queratohial |
1 |
0 |
0 |
1 |
1 |
2,4 |
13000 |
||||||
| pII-5 |
Actinopterygii |
Ariidae |
Selenaspsis |
dowii |
queratohial |
0 |
1 |
0 |
1 |
1 |
4,1 |
13000 |
||||||
| pII-4 |
Actinopterygii |
Ariidae |
Selenaspsis |
dowii |
pteriogóforo dorsal, I |
0 |
0 |
0 |
1 |
1 |
6,6 |
13000 |
||||||
| pII-4 |
Actinopterygii |
Ariidae |
Selenaspsis |
dowii |
urohial |
0 |
0 |
0 |
1 |
1 |
1 |
13000 |
||||||
| pII-3 |
Actinopterygii |
Ariidae |
Selenaspsis |
dowii |
complejo vert. fusionado |
0 |
0 |
0 |
1 |
1 |
6,6 |
11000 |
||||||
| pIII-1 |
Actinopterygii |
Ariidae |
Selenaspsis |
dowii |
esfenótico |
0 |
1 |
0 |
1 |
1 |
6,5 |
11000 |
||||||
| A22 II NA |
Actinopterygii |
Ariidae |
Selenaspsis |
dowii |
esfenótico |
0 |
1 |
0 |
1 |
1 |
4,2 |
11000 |
2 |
22000 |
||||
| A 22 CII DT 2 p II-8 |
Actinopterygii |
Ariidae |
Selenaspsis |
dowii |
espina dorsal |
0 |
0 |
0 |
1 |
1 |
4,9 |
11000 |
||||||
| pII-12 |
Actinopterygii |
Ariidae |
Selenaspsis |
dowii |
opérculo |
1 |
0 |
0 |
1 |
1 |
3,1 |
11000 |
||||||
| pII-3 |
Actinopterygii |
Ariidae |
Selenaspsis |
dowii |
preopérculo |
0 |
1 |
0 |
1 |
1 |
0,7 |
11000 |
||||||
| pII-3 |
Actinopterygii |
Ariidae |
Selenaspsis |
dowii |
supraoccipital |
0 |
0 |
0 |
1 |
1 |
3,3 |
11000 |
||||||
| pII-3 |
Actinopterygii |
Ariidae |
Selenaspsis |
dowii |
pteriogóforo dorsal, I |
0 |
0 |
0 |
1 |
1 |
0,6 |
11000 |
||||||
| pII-2 |
Actinopterygii |
Ariidae |
Selenaspsis |
dowii |
pterótico |
1 |
0 |
0 |
1 |
1 |
4,1 |
11000 |
||||||
| pII-3 |
Actinopterygii |
Ariidae |
Selenaspsis |
dowii |
vértebra |
0 |
0 |
0 |
5 |
5 |
5 |
11000 |
||||||
| pIII-2 |
Actinopterygii |
Ariidae |
Selenaspsis |
dowii |
faringeal |
1 |
0 |
0 |
1 |
1 |
1 |
9000 |
||||||
| pIII-2 |
Actinopterygii |
Ariidae |
Selenaspsis |
dowii |
supraoccipital |
0 |
0 |
0 |
1 |
1 |
1,3 |
9000 |
||||||
| pIII-2 |
Actinopterygii |
Ariidae |
Selenaspsis |
dowii |
pteriogóforo dorsal, I |
0 |
0 |
0 |
1 |
1 |
0,5 |
9000 |
||||||
| pIII-2 |
Actinopterygii |
Ariidae |
Selenaspsis |
dowii |
vértebra |
0 |
0 |
0 |
1 |
1 |
0,5 |
9000 |
1 |
9000 |
||||
| pII-2 |
Actinopterygii |
Ariidae |
Selenaspsis |
dowii |
cleitron |
1 |
0 |
0 |
1 |
1 |
1,5 |
4000 |
||||||
| pII-4 |
Actinopterygii |
Ariidae |
Selenaspsis |
dowii |
cleitron |
0 |
1 |
0 |
1 |
1 |
4,3 |
4000 |
||||||
| A22 Cii AT2 pII-8 |
Actinopterygii |
Ariidae |
Selenaspsis |
dowii |
complejo vert. fusionado |
0 |
0 |
0 |
1 |
1 |
2,2 |
4000 |
||||||
| pII-2 |
Actinopterygii |
Ariidae |
Selenaspsis |
dowii |
complejo vert. fusionado |
0 |
0 |
0 |
1 |
1 |
2,2 |
4000 |
||||||
| pII-2 |
Actinopterygii |
Ariidae |
Selenaspsis |
dowii |
esfenótico |
1 |
0 |
0 |
1 |
1 |
2,8 |
4000 |
||||||
| A22 Cii AT2 pII-8 |
Actinopterygii |
Ariidae |
Selenaspsis |
dowii |
esfenótico |
1 |
0 |
0 |
1 |
1 |
1,5 |
4000 |
||||||
| pII-5 |
Actinopterygii |
Ariidae |
Selenaspsis |
dowii |
espina dorsal |
0 |
0 |
0 |
1 |
1 |
3,6 |
4000 |
||||||
| pIII-1 |
Actinopterygii |
Ariidae |
Selenaspsis |
dowii |
fragmento |
0 |
0 |
0 |
1 |
1 |
1,1 |
4000 |
||||||
| pII-2 |
Actinopterygii |
Ariidae |
Selenaspsis |
dowii |
frontal |
0 |
1 |
0 |
1 |
1 |
5,2 |
4000 |
||||||
| A22 C22 AT2 |
Actinopterygii |
Ariidae |
Selenaspsis |
dowii |
frontal |
0 |
1 |
0 |
1 |
1 |
1,3 |
4000 |
2 |
8000 |
||||
| p-III |
Actinopterygii |
Ariidae |
Selenaspsis |
dowii |
hipohial |
1 |
0 |
0 |
1 |
1 |
0,5 |
4000 |
||||||
| pII-2 |
Actinopterygii |
Ariidae |
Selenaspsis |
dowii |
post-temporal |
1 |
0 |
0 |
1 |
1 |
1,5 |
4000 |
||||||
| pII-3 |
Actinopterygii |
Ariidae |
Selenaspsis |
dowii |
prootic |
0 |
1 |
0 |
1 |
1 |
1,2 |
4000 |
||||||
| pII-3 |
Actinopterygii |
Ariidae |
Selenaspsis |
dowii |
espina dorsal |
0 |
0 |
0 |
1 |
1 |
1,1 |
2000 |
1 |
2000 |
||||
| pIII-2 |
Actinopterygii |
Belonidae |
Tylosurus |
cocodrilus |
vértebra |
0 |
0 |
0 |
4 |
4 |
0,7 |
700 |
1 |
700 |
||||
| pII-3 |
Actinopterygii |
Carangidae |
Caranx |
vértebra |
0 |
0 |
0 |
1 |
1 |
0,3 |
1400 |
1 |
1400 |
|||||
| pIII-2 |
Actinopterygii |
Carangidae |
Caranx |
caballus |
dentario |
0 |
1 |
0 |
1 |
1 |
0,1 |
400 |
||||||
| pIII-2 |
Actinopterygii |
Carangidae |
Caranx |
caballus |
vértebra |
0 |
0 |
0 |
1 |
1 |
0,2 |
400 |
1 |
400 |
||||
| pIII-2 |
Actinopterygii |
Carangidae |
Caranx |
caballus |
premaxila |
1 |
0 |
0 |
1 |
1 |
0,1 |
325 |
||||||
| pIII-2 |
Actinopterygii |
Carangidae |
Caranx |
caballus |
vértebra |
5 |
0 |
0 |
0 |
1 |
1 |
0,2 |
325 |
|||||
| pIII-2 |
Actinopterygii |
Carangidae |
Caranx |
caballus |
vértebra |
6 |
0 |
0 |
0 |
1 |
1 |
0,1 |
325 |
|||||
| pIII-2 |
Actinopterygii |
Carangidae |
Caranx |
caballus |
vértebra |
6 |
0 |
0 |
0 |
1 |
1 |
0,1 |
325 |
2 |
650 |
|||
| pIII-2 |
Actinopterygii |
Carangidae |
Caranx |
caballus |
vértebra |
9 |
0 |
0 |
0 |
1 |
1 |
0,2 |
325 |
|||||
| pIII-2 |
Actinopterygii |
Carangidae |
Caranx |
caballus |
frontal |
0 |
1 |
0 |
1 |
1 |
0,1 |
250 |
||||||
| pIII-2 |
Actinopterygii |
Carangidae |
Caranx |
caballus |
opérculo |
1 |
0 |
0 |
1 |
1 |
0,1 |
250 |
1 |
250 |
||||
| pII-3 |
Actinopterygii |
Carangidae |
Caranx |
caninus |
maxila |
1 |
0 |
0 |
1 |
1 |
0,2 |
650 |
||||||
| pIII-2 |
Actinopterygii |
Carangidae |
Caranx |
caninus |
vértebra |
0 |
0 |
0 |
1 |
1 |
0,3 |
650 |
1 |
650 |
||||
| pII-3 |
Actinopterygii |
Carangidae |
Caranx |
caninus |
maxila |
0 |
1 |
0 |
1 |
1 |
0,1 |
400 |
1 |
400 |
||||
| pIII-2 |
Actinopterygii |
Carangidae |
Caranx |
caninus |
vértebra |
0 |
0 |
0 |
1 |
1 |
0,1 |
100 |
1 |
100 |
||||
| pII-2 |
Actinopterygii |
Carangidae |
Caranx |
speciosus |
hiomandíbula |
1 |
0 |
0 |
1 |
1 |
1,6 |
3200 |
||||||
| pII-5 |
Actinopterygii |
Carangidae |
Caranx |
speciosus |
vértebra |
7 |
0 |
0 |
0 |
1 |
1 |
1,3 |
3200 |
|||||
| pII-2 |
Actinopterygii |
Carangidae |
Caranx |
speciosus |
hiomandíbula |
9 |
1 |
0 |
0 |
1 |
1 |
1,8 |
3200 |
1 |
3200 |
|||
| pIII-2 |
Actinopterygii |
Carangidae |
Chloroscombrus |
orqueta |
cleitron |
0 |
1 |
0 |
1 |
1 |
0,1 |
40 |
1 |
40 |
||||
| pIII-2 |
Actinopterygii |
Carangidae |
Hemicaranx |
leucurus |
opérculo |
0 |
1 |
0 |
1 |
1 |
0,1 |
300 |
1 |
300 |
||||
| A22 II PA |
Actinopterygii |
Carangidae |
Oligoplites |
altus |
vértebra |
22 |
0 |
0 |
0 |
1 |
1 |
0,6 |
1000 |
|||||
| pIII-2 |
Actinopterygii |
Carangidae |
Oligoplites |
altus |
vértebra |
0 |
0 |
0 |
1 |
1 |
0,2 |
1000 |
1 |
1000 |
||||
| pII-3 |
Actinopterygii |
Carangidae |
Oligoplites |
altus |
vértebra |
0 |
0 |
0 |
3 |
3 |
1 |
700 |
1 |
700 |
||||
| pIII-2 |
Actinopterygii |
Carangidae |
Selene |
oerstedii |
vértebra |
0 |
0 |
0 |
1 |
1 |
0,1 |
500 |
1 |
500 |
||||
| pIII-2 |
Actinopterygii |
Carangidae |
Selene |
peruviana |
hiomandíbula |
1 |
0 |
0 |
1 |
1 |
0,1 |
150 |
||||||
| pIII-2 |
Actinopterygii |
Carangidae |
Selene |
peruviana |
premaxila |
1 |
0 |
0 |
1 |
1 |
0,1 |
150 |
||||||
| pIII-2 |
Actinopterygii |
Carangidae |
Selene |
peruviana |
vértebra |
5 |
0 |
0 |
0 |
1 |
1 |
0,1 |
150 |
1 |
150 |
|||
| pIII-2 |
Actinopterygii |
Carangidae |
Selene |
peruviana |
premaxila |
0 |
1 |
0 |
1 |
1 |
0,1 |
100 |
||||||
| pIII-2 |
Actinopterygii |
Carangidae |
Selene |
peruviana |
pteriogóforo |
0 |
0 |
0 |
1 |
1 |
0,1 |
100 |
1 |
100 |
||||
| pII-2 |
Actinopterygii |
Centropomidae |
Centropomus |
espina anal |
0 |
0 |
0 |
1 |
1 |
1,6 |
||||||||
| pII-3 |
Actinopterygii |
Centropomidae |
Centropomus |
espina anal |
0 |
0 |
0 |
1 |
1 |
1,9 |
||||||||
| pIII-1 |
Actinopterygii |
Centropomidae |
Centropomus |
espina anal |
0 |
0 |
0 |
1 |
1 |
0,4 |
||||||||
| pII-3 |
Actinopterygii |
Centropomidae |
Centropomus |
espina |
0 |
0 |
0 |
1 |
1 |
0,4 |
||||||||
| pII-3 |
Actinopterygii |
Centropomidae |
Centropomus |
vértebra |
0 |
0 |
0 |
1 |
1 |
0,2 |
||||||||
| pII-3 |
Actinopterygii |
Centropomidae |
Centropomus |
vértebra |
0 |
0 |
0 |
3 |
3 |
0,4 |
||||||||
| pII-3 |
Actinopterygii |
Centropomidae |
Centropomus |
armatus |
espina anal |
0 |
0 |
0 |
1 |
1 |
3,7 |
1500 |
1 |
1500 |
||||
| pIII-2 |
Actinopterygii |
Centropomidae |
Centropomus |
armatus |
espina anal |
0 |
0 |
0 |
1 |
1 |
3,8 |
1200 |
1 |
1200 |
||||
| pII-2 |
Actinopterygii |
Centropomidae |
Centropomus |
armatus |
espina anal |
0 |
0 |
0 |
1 |
1 |
0,9 |
500 |
||||||
| pII-3 |
Actinopterygii |
Centropomidae |
Centropomus |
armatus |
vértebra |
4 |
0 |
0 |
0 |
1 |
1 |
0,5 |
500 |
1 |
500 |
|||
| pII-5 |
Actinopterygii |
Centropomidae |
Centropomus |
armatus |
vértebra |
0 |
0 |
0 |
1 |
1 |
1 |
200 |
1 |
200 |
||||
| pII-5 |
Actinopterygii |
Centropomidae |
Centropomus |
medius |
vértebra |
1 |
0 |
0 |
0 |
1 |
1 |
1,1 |
2300 |
|||||
| pII-5 |
Actinopterygii |
Centropomidae |
Centropomus |
medius |
vértebra |
7 |
0 |
0 |
0 |
1 |
1 |
2 |
2300 |
|||||
| pII-3 |
Actinopterygii |
Centropomidae |
Centropomus |
medius |
vértebra |
3 |
0 |
0 |
0 |
1 |
1 |
0,4 |
2300 |
1 |
2300 |
|||
| pII-3 |
Actinopterygii |
Centropomidae |
Centropomus |
medius |
vértebra |
10 |
0 |
0 |
0 |
1 |
1 |
1,1 |
1300 |
1 |
1300 |
|||
| pII-3 |
Actinopterygii |
Centropomidae |
Centropomus |
medius |
vértebra |
18 |
0 |
0 |
0 |
1 |
1 |
0,6 |
1000 |
|||||
| pII-3 |
Actinopterygii |
Centropomidae |
Centropomus |
medius |
vértebra |
21 |
0 |
0 |
0 |
1 |
1 |
0,7 |
1000 |
1 |
1000 |
|||
| pII-3 |
Actinopterygii |
Centropomidae |
Centropomus |
medius |
espina anal |
0 |
0 |
0 |
1 |
1 |
0,7 |
300 |
1 |
300 |
||||
| pII-2 |
Actinopterygii |
Centropomidae |
Centropomus |
nigrescens |
queratohial |
1 |
0 |
0 |
1 |
1 |
2 |
5000 |
||||||
| pII-2 |
Actinopterygii |
Centropomidae |
Centropomus |
nigrescens |
vértebra |
6 |
0 |
0 |
0 |
1 |
1 |
3,2 |
5000 |
1 |
5000 |
|||
| pIII-1 |
Actinopterygii |
Centropomidae |
Centropomus |
nigrescens |
vértebra |
5 |
0 |
0 |
0 |
1 |
1 |
2,1 |
3200 |
1 |
3200 |
|||
| pIII-2 |
Actinopterygii |
Centropomidae |
Centropomus |
nigrescens |
vértebra |
1 |
0 |
0 |
0 |
1 |
1 |
0,6 |
1000 |
1 |
1000 |
|||
| pIII-2 |
Actinopterygii |
Centropomidae |
Centropomus |
viridis |
vértebra |
2 |
0 |
0 |
0 |
1 |
1 |
2,2 |
4000 |
1 |
4000 |
|||
| A22 CII At2 pII-8 |
Actinopterygii |
Centropomidae |
Centropomus |
viridis |
vértebra |
1 |
0 |
0 |
0 |
1 |
1 |
1,5 |
2300 |
|||||
| pII-4 |
Actinopterygii |
Centropomidae |
Centropomus |
viridis |
vértebra |
6 |
0 |
0 |
0 |
1 |
1 |
1,8 |
2300 |
|||||
| pII-4 |
Actinopterygii |
Centropomidae |
Centropomus |
viridis |
vértebra |
8 |
0 |
0 |
0 |
1 |
1 |
2,1 |
2300 |
1 |
2300 |
|||
| pII-3 |
Actinopterygii |
Centropomidae |
Centropomus |
viridis |
vértebra |
6 |
0 |
0 |
0 |
1 |
1 |
0,7 |
900 |
1 |
900 |
|||
| pIII-2 |
Actinopterygii |
Chaetodontidae |
Chaetodon |
humeralis |
vértebra |
9 |
0 |
0 |
0 |
1 |
1 |
0,05 |
60 |
1 |
60 |
|||
| pIII-2 |
Actinopterygii |
Clupeidae |
Opisthonema |
cf |
libertate |
proótico |
0 |
1 |
1 |
0 |
1 |
0,01 |
150 |
1 |
150 |
|||
| pIII-2 |
Actinopterygii |
Gerreidae |
Eucinostomus |
currani |
vértebra |
4 |
0 |
0 |
0 |
1 |
1 |
0,06 |
80 |
1 |
80 |
|||
| pIII-2 |
Actinopterygii |
Gobiidae |
Bathygobius |
vértebra |
0 |
0 |
0 |
1 |
1 |
0,05 |
100 |
1 |
100 |
|||||
| pIII-2 |
Actinopterygii |
Haemulidae |
Anisotremus |
taeniatus |
cuadrado |
1 |
0 |
0 |
1 |
1 |
0,05 |
125 |
1 |
125 |
||||
| pIII-2 |
Actinopterygii |
Haemulidae |
Haemulopsis |
elongatus |
hiomandíbula |
0 |
1 |
0 |
1 |
1 |
0,2 |
250 |
1 |
250 |
||||
| pIII-2 |
Actinopterygii |
Haemulidae |
Haemulopsis |
elongatus |
hiomandíbula |
0 |
1 |
0 |
1 |
1 |
0,08 |
125 |
1 |
125 |
||||
| pIII-2 |
Actinopterygii |
Haemulidae |
Haemulopsis |
leuciscus |
vértebra |
22 |
0 |
0 |
0 |
1 |
1 |
0,2 |
325 |
1 |
325 |
|||
| pIII-2 |
Actinopterygii |
Haemulidae |
Orthopristis |
chalceus |
pteriogóforo anal 1 |
0 |
0 |
0 |
3 |
3 |
0,1 |
300 |
3 |
900 |
||||
| pIII-2 |
Actinopterygii |
Haemulidae |
Orthopristis |
chalceus |
vértebra |
1 |
0 |
0 |
0 |
1 |
1 |
0,08 |
300 |
|||||
| pIII-2 |
Actinopterygii |
Haemulidae |
Orthopristis |
chalceus |
cuadrado |
1 |
0 |
0 |
1 |
1 |
0,04 |
275 |
||||||
| pIII-2 |
Actinopterygii |
Haemulidae |
Orthopristis |
chalceus |
hiomandíbula |
0 |
1 |
0 |
1 |
1 |
0,07 |
275 |
||||||
| pIII-2 |
Actinopterygii |
Haemulidae |
Orthopristis |
chalceus |
post-temporal |
0 |
1 |
0 |
1 |
1 |
0,05 |
275 |
||||||
| pIII-2 |
Actinopterygii |
Haemulidae |
Orthopristis |
chalceus |
supraoccipital |
0 |
0 |
0 |
1 |
1 |
0,08 |
275 |
||||||
| pIII-2 |
Actinopterygii |
Haemulidae |
Orthopristis |
chalceus |
pteriogóforo anal 1 |
0 |
0 |
0 |
2 |
2 |
0,1 |
275 |
2 |
550 |
||||
| pIII-2 |
Actinopterygii |
Haemulidae |
Orthopristis |
chalceus |
vértebra |
7 |
0 |
0 |
0 |
2 |
2 |
0,1 |
275 |
|||||
| pIII-2 |
Actinopterygii |
Haemulidae |
Orthopristis |
chalceus |
vértebra |
8 |
0 |
0 |
0 |
1 |
1 |
0,07 |
275 |
|||||
| pIII-2 |
Actinopterygii |
Haemulidae |
Orthopristis |
chalceus |
vértebra |
0 |
0 |
0 |
5 |
5 |
0,2 |
275 |
||||||
| pIII-2 |
Actinopterygii |
Haemulidae |
Orthopristis |
chalceus |
epihial |
0 |
1 |
0 |
1 |
1 |
0,03 |
175 |
||||||
| pIII-2 |
Actinopterygii |
Haemulidae |
Orthopristis |
chalceus |
parasphenoid |
0 |
0 |
0 |
1 |
1 |
0,09 |
175 |
||||||
| pIII-2 |
Actinopterygii |
Haemulidae |
Orthopristis |
chalceus |
vértebra |
1 |
0 |
0 |
0 |
1 |
1 |
0,07 |
175 |
|||||
| pIII-2 |
Actinopterygii |
Haemulidae |
Orthopristis |
chalceus |
vómer |
0 |
0 |
0 |
1 |
1 |
0,06 |
175 |
1 |
175 |
||||
| pIII-2 |
Actinopterygii |
Haemulidae |
Orthopristis |
chalceus |
cleitron |
0 |
1 |
0 |
1 |
1 |
0,05 |
60 |
1 |
60 |
||||
| pIII-2 |
Actinopterygii |
Haemulidae |
Pomadasys |
macracanthus |
vértebra |
0 |
0 |
0 |
1 |
1 |
0,2 |
650 |
1 |
650 |
||||
| pIII-2 |
Actinopterygii |
Haemulidae |
Pomadasys |
panamensis |
vértebra |
8 |
0 |
0 |
0 |
1 |
1 |
0,1 |
350 |
1 |
350 |
|||
| pIII-2 |
Actinopterygii |
Kyphosidae |
Kyphosus |
vértebra |
0 |
0 |
0 |
1 |
1 |
0,1 |
700 |
1 |
700 |
|||||
| pIII-2 |
Actinopterygii |
Kyphosidae |
Sectator |
ocyurus |
vértebra |
0 |
0 |
0 |
2 |
2 |
0,1 |
400 |
1 |
400 |
||||
| pIII-2 |
Actinopterygii |
Lobotidae |
Lobotes |
surinamensis |
dentario |
0 |
1 |
0 |
1 |
1 |
6,7 |
5300 |
||||||
| pIII-2 |
Actinopterygii |
Lobotidae |
Lobotes |
surinamensis |
espina |
0 |
0 |
0 |
1 |
1 |
1,1 |
5300 |
||||||
| pII-2 |
Actinopterygii |
Lobotidae |
Lobotes |
surinamensis |
vértebra |
5 |
0 |
0 |
0 |
1 |
1 |
3,3 |
5300 |
|||||
| pII-4 |
Actinopterygii |
Lobotidae |
Lobotes |
surinamensis |
vértebra |
0 |
0 |
0 |
1 |
1 |
2 |
5300 |
1 |
5300 |
||||
| pII-3 |
Actinopterygii |
Lobotidae |
Lobotes |
surinamensis |
vértebra |
9 |
0 |
0 |
0 |
1 |
1 |
2,2 |
3700 |
|||||
| pII-3 |
Actinopterygii |
Lobotidae |
Lobotes |
surinamensis |
vértebra |
12 |
0 |
0 |
0 |
1 |
1 |
2,4 |
3700 |
1 |
3700 |
|||
| pII-2 |
Actinopterygii |
Lobotidae |
Lobotes |
surinamensis |
articular |
0 |
1 |
0 |
1 |
1 |
2,4 |
2500 |
||||||
| pII-6 |
Actinopterygii |
Lobotidae |
Lobotes |
surinamensis |
vértebra |
9 |
0 |
0 |
0 |
1 |
1 |
2,4 |
2500 |
1 |
2500 |
|||
| pII-2 |
Actinopterygii |
Lobotidae |
Lobotes |
surinamensis |
vértebra |
3 |
0 |
0 |
0 |
1 |
1 |
1,3 |
2000 |
|||||
| pII-2 |
Actinopterygii |
Lobotidae |
Lobotes |
surinamensis |
vértebra |
5 |
0 |
0 |
0 |
1 |
1 |
0,09 |
2000 |
1 |
2000 |
|||
| pII-5 |
Actinopterygii |
Lobotidae |
Lobotes |
surinamensis |
cleitron |
0 |
1 |
0 |
1 |
1 |
0,7 |
1500 |
||||||
| pII-3 |
Actinopterygii |
Lobotidae |
Lobotes |
surinamensis |
vértebra |
0 |
0 |
0 |
3 |
3 |
1,6 |
1500 |
1 |
1500 |
||||
| pIII-2 |
Actinopterygii |
Lutjanidae |
Lutjanus |
guttatus |
vértebra |
0 |
0 |
0 |
2 |
2 |
2 |
2500 |
1 |
2500 |
||||
| pIII-2 |
Actinopterygii |
Mugilidae |
Chaenomugil |
proboscidens |
vértebra |
0 |
0 |
0 |
2 |
2 |
0,09 |
125 |
1 |
125 |
||||
| pIII-2 |
Actinopterygii |
Mugilidae |
Mugil |
cf |
curema4 |
vértebra |
0 |
0 |
0 |
2 |
2 |
0,4 |
450 |
1 |
450 |
|||
| pIII-2 |
Actinopterygii |
Mugilidae |
Mugil |
cf |
curema |
vértebra |
0 |
0 |
0 |
1 |
1 |
0,08 |
150 |
1 |
150 |
|||
| pIII-2 |
Actinopterygii |
Polynemidae |
Polydactylus |
opercularis |
vértebra |
22 |
0 |
0 |
0 |
1 |
1 |
0,07 |
250 |
1 |
250 |
|||
| pIII-2 |
Actinopterygii |
Pristigasteridae |
Ilisha |
furthii |
vértebra |
0 |
0 |
0 |
3 |
3 |
0,1 |
250 |
1 |
250 |
||||
| pIII-2 |
Actinopterygii |
Sciaenidae |
Bairdiella |
armata |
queratohial |
1 |
0 |
0 |
1 |
1 |
0,08 |
225 |
1 |
225 |
||||
| pIII-2 |
Actinopterygii |
Sciaenidae |
Bairdiella |
armata |
queratohial |
0 |
1 |
0 |
1 |
1 |
0,06 |
50 |
1 |
50 |
||||
| pII-3 |
Actinopterygii |
Sciaenidae |
Cynoscion |
cf |
albus |
vértebra |
0 |
0 |
0 |
1 |
1 |
0,6 |
1400 |
1 |
1400 |
|||
| pIII-2 |
Actinopterygii |
Sciaenidae |
Cynoscion |
albus |
cleitron |
0 |
1 |
0 |
1 |
1 |
2,1 |
4500 |
1 |
4500 |
||||
| pIII-2 |
Actinopterygii |
Sciaenidae |
Cynoscion |
albus |
vértebra |
0 |
0 |
0 |
1 |
1 |
1,6 |
2200 |
1 |
2200 |
||||
| pIII-2 |
Actinopterygii |
Sciaenidae |
Cynoscion |
albus |
vértebra |
0 |
0 |
0 |
1 |
1 |
0,08 |
200 |
1 |
200 |
||||
| pIII-2 |
Actinopterygii |
Sciaenidae |
Cynoscion |
squamipinnis |
hiomandíbula |
0 |
1 |
0 |
1 |
1 |
0,5 |
2300 |
1 |
2300 |
||||
| pIII-2 |
Actinopterygii |
Sciaenidae |
Larimus |
acclivis |
vértebra |
5 |
0 |
0 |
0 |
1 |
1 |
0,09 |
100 |
1 |
100 |
|||
| pIII-2 |
Actinopterygii |
Sciaenidae |
Larimus |
acclivis |
vértebra |
0 |
0 |
0 |
2 |
2 |
0,3 |
450 |
1 |
450 |
||||
| pIII-2 |
Actinopterygii |
Sciaenidae |
Micropogonias |
altipinnis |
cuadrado |
1 |
0 |
0 |
1 |
1 |
0,09 |
150 |
||||||
| pIII-2 |
Actinopterygii |
Sciaenidae |
Micropogonias |
altipinnis |
queratohial |
0 |
1 |
0 |
1 |
1 |
0,08 |
150 |
||||||
| pIII-2 |
Actinopterygii |
Sciaenidae |
Micropogonias |
altipinnis |
vértebra |
0 |
0 |
0 |
2 |
2 |
0,3 |
150 |
1 |
150 |
||||
| pIII-2 |
Actinopterygii |
Sciaenidae |
Ophioscion |
cf |
vermicularis |
pteriogóforo anal I |
0 |
0 |
0 |
1 |
1 |
0,1 |
125 |
1 |
125 |
|||
| pIII-2 |
Actinopterygii |
Sciaenidae |
Paralonchurus |
petersi |
vértebra |
8 |
0 |
0 |
0 |
1 |
1 |
0,06 |
40 |
1 |
40 |
|||
| pIII-2 |
Actinopterygii |
Sciaenidae |
Stellifer |
chrysoleuca |
vértebra |
0 |
0 |
0 |
2 |
2 |
0,3 |
350 |
1 |
350 |
||||
| pIII-2 |
Actinopterygii |
Sciaenidae |
Stellifer |
furthii |
pteriogóforo anal I |
0 |
0 |
0 |
1 |
1 |
0,08 |
70 |
1 |
70 |
||||
| pIII-2 |
Actinopterygii |
Sciaenidae |
Stellifer |
furthii |
pteriogóforo anal I |
0 |
0 |
0 |
1 |
1 |
0,08 |
50 |
1 |
50 |
||||
| pII-3 |
Actinopterygii |
cf |
Serranidae |
paresfenoides |
0 |
0 |
0 |
1 |
1 |
9,8 |
||||||||
| pII-3 |
Actinopterygii |
Serranidae |
Epinephelus |
espina dorsal |
0 |
0 |
0 |
1 |
1 |
5,1 |
40.000 |
|||||||
| pII-5 |
Actinopterygii |
Serranidae |
Epinephelus |
espina dorsal |
0 |
0 |
0 |
1 |
1 |
7,9 |
40.000 |
1 |
40.000 |
|||||
| pIII-2 |
Actinopterygii |
Sphyraenidae |
Sphyraena |
cf |
ensis |
maxila |
0 |
1 |
0 |
1 |
1 |
0,1 |
400 |
|||||
| pIII-2 |
Actinopterygii |
Sphyraenidae |
Sphyraena |
cf |
ensis |
vértebra |
0 |
0 |
0 |
2 |
2 |
0,3 |
400 |
1 |
400 |
|||
| 33 |
38 |
1 |
263 |
264 |
236,91 |
100 |
198405 |
| Notas: 1 La morfología del lapilo (otolito grande) y de los huesos del neurocráneo sugieren que este género es politípico (ver Cooke 1996) |
|
| 2 Aunque esta especie etá asignada por lo general al género Sciadeops (cf. Allen y Robertson 1994), su lapilo se asemeja al de muchas especies atribuidas a Arius por lo que se cuestiona su colocación en un género monotípico |
|
| 3 Esta especie diverge en muchos aspectos de los demás aríidos del Pacífico tropical oriental. Existe bastante confusión en la literatura ictiológica en torno a la prioridad del nombre genérico; algunos autores la asignan a Sciadeichthys (cf Allen y Robertson |
|
| 1994) o Arius (cf Castro-Aguirre 1978:49). |
|
| 4 No poseemos especímenes de Mugil cephalus, especie que nunca hemos colectado en Panamá aunque se supone que debería de frecuentar aguas estuarinas istmeñas. |
|
Cuadro 4
| Familia |
Nombre |
Núm de géneros |
Núm de especies |
Núm de especies tentativas |
Núm de huesos |
% de huesos |
Núm mínimo de individuos |
% NMI |
Biomasa |
% biomasa |
|
| 1 |
Carcharhinidae |
tiburones |
3 |
2 |
12 |
5,5 |
3 |
3 |
8600 |
4,4 |
|
| 2 |
Sphyrnidae |
tiburones martillo |
1 |
1 |
1 |
0,45 |
1 |
1 |
700 |
0,4 |
|
| 3 |
Ariidae |
bagres, cominates y congos |
4 |
6 |
1 |
64 |
29,1 |
18 |
18 |
77530 |
39 |
| 4 |
Belonidae |
peces aguja |
1 |
1 |
1 |
0,45 |
1 |
1 |
700 |
0,4 |
|
| 5 |
Carangidae |
jureles, pómpanos |
5 |
8 |
30 |
13,6 |
16 |
16 |
9840 |
5 |
|
| 6 |
Centropomidae |
robalos |
1 |
4 |
29 |
13,2 |
14 |
14 |
24700 |
13 |
|
| 7 |
Chaetodontidae |
peces mariposa |
1 |
1 |
1 |
0,45 |
1 |
1 |
60 |
0 |
|
| 8 |
Clupeidae |
arenques |
1 |
1 |
1 |
0,45 |
1 |
1 |
150 |
0,1 |
|
| 9 |
Gerreidae |
mojarras marinas |
1 |
1 |
1 |
0,45 |
1 |
1 |
80 |
0 |
|
| 10 |
Gobiidae |
gobíidos |
1 |
1 |
0,45 |
1 |
1 |
100 |
0,1 |
||
| 11 |
Hamulidae |
roncadores |
4 |
6 |
29 |
13,2 |
13 |
13 |
3510 |
1,8 |
|
| 12 |
Kyphosidae |
chopas |
2 |
1 |
3 |
1,4 |
2 |
2 |
1100 |
0,6 |
|
| 13 |
Lobotidae |
berrugates |
1 |
1 |
14 |
6,4 |
5 |
5 |
15000 |
7,6 |
|
| 14 |
Lutjanidae |
pargos |
1 |
1 |
2 |
0,9 |
1 |
1 |
1500 |
0,8 |
|
| 15 |
Mugilidae |
lisas |
2 |
1 |
1 |
5 |
2,3 |
3 |
3 |
725 |
0,4 |
| 16 |
Polynemidae |
bobos |
1 |
1 |
1 |
0,45 |
1 |
1 |
250 |
0,1 |
|
| 17 |
Pristigasteridae |
machetes |
1 |
1 |
1 |
0,45 |
1 |
1 |
250 |
0,1 |
|
| 18 |
Sciaenidae |
corvinas y coscochas |
7 |
8 |
1 |
19 |
8,6 |
15 |
15 |
12210 |
6,2 |
| 19 |
Serranidae |
meros y cabrillas |
1 |
2 |
0,9 |
1 |
1 |
40000 |
20 |
||
| 20 |
Sphyraenidae |
barracudas |
1 |
1 |
3 |
1,4 |
1 |
1 |
400 |
0 |
|
| TOTAL: |
40 |
43 |
6 |
220 |
100 |
100 |
100 |
197405 |
100 |
Cuadro 5
Cuadro 6
| Estuario alto y medio (incluidos manglares) |
Estuario bajo (margen externo de la zona de mezcolanza) |
Aguas someras libres de sedimentos suspendidos (p.ejm., playas, bancos de arena y corrientes de aguas claras en estuarios) |
Arrecifes y rocas (incluidos islas y farallones) |
| Carcharhinus leucas |
Carcharhinus leucas |
Rhizoprionodon longurio |
TYLOSURUS COCODRILUS |
| Rhizoprionodon longurio |
Rhizoprionodon longurio |
Tylosurus cocodrilus |
Caranx caballus |
| Sphyrna cf lewini |
Arius dasycephalus |
Caranx caballus |
CARANX CANINUS |
| Bagre pinnimaculatus |
Arius troschelii |
Caranx caninus |
Caranx speciosus |
| Cathorops hypophthalmus |
Bagre pinnimaculatus |
Caranx speciosus |
OLIGOPLITES ALTUS |
| Cathorops multiradiatus |
Cathorops multiradiatus |
Chloroscombrus orqueta |
SELENE OERSTEDII |
| Cathorops tuyra |
Caranx caballus |
Hemicaranx leucurus |
Chaetodon humeralis |
| Caranx caninus |
Hemicaranx leucurus |
Selene oerstedii |
Kyphosus |
| Chloroscombrus orqueta |
Cathorops multiradiatus |
Selene peruviana |
Sectator ocyurus |
| Hemicaranx leucurus |
Tylosurus cocodrilus |
Opisthonema |
LUTJANUS GUTTATUS |
| Oligoplites altus |
Caranx caballus |
Eucinostomus currani |
Epinephelus |
| Selene oerstedii |
Caranx caninus |
Haemulopsis elongatus |
SPHYRAENA ENSIS |
| Centropomus armatus |
Hemicaranx leucurus |
Haemulopsis leuciscus |
|
| Centropomus medius |
Selene peruviana |
Orthopristis chalceus |
|
| Centropomus nigrescens |
Opisthonema |
Pomadasys panamensis |
|
| Centropomus viridis |
HAEMULOPSIS ELONGATUS |
Lutjanus guttatus |
|
| Eucinostomus currani |
HAEMULOPSIS LEUCISCUS |
Chaenomugil probodiscens |
|
| Bathygobius |
Orthopristis chalceus |
Mugil curema |
|
| Haemulopsis elongatus |
Pomadasys panamensis |
Polydactylus opercularis |
|
| Haemulopsis leuciscus |
Lutjanus guttatus |
Larimus acclivis |
|
| Pomadasys macracanthus |
Cynoscion squamipinnis |
Epinephelus |
|
| Lobotes surinamensis |
MICROPOGONIAS ALTIPINNIS |
Sphyraena ensis |
|
| Mugil curema |
Paralonchurus petersi |
||
| Polydactylus opercularis |
Epinephelus |
||
| Ilisha furthii |
|||
| Bairdiella armata |
|||
| Cynoscion albus |
|||
| Larimus acclivis |
|||
| Micropogonias altipinnis |
|||
| Ophioscion vermicularis |
|||
| Stellifer chrysoleuca |
|||
| Stellifer furthii |
|||
| Epinephelus |
Leyenda:
Bairdiella armata Hábitat preferido
Micropogonias altipinnis Prefiere este habitat cuando en estado juvenil, sub-adulto o adulto pequeño
TYLOSURUS COCODRILUS Se encuentra en este hábitat con mayor frecuencia cuando es adulto maduro
Cuadro 7
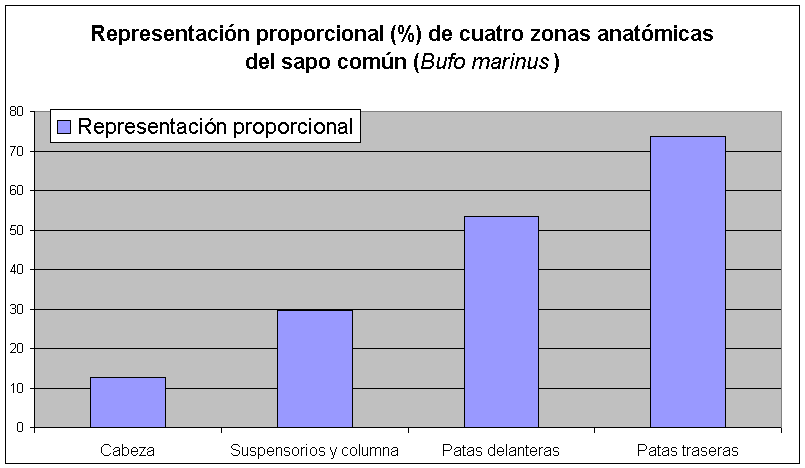
BIBLIOGRAFÍA
Allen, Gerald R. y D. Ross Robertson
1994 The complete divers and fishermens guide to fishes of the tropical eastern Pacific. Crawford House Press, Bathurst.
Castro-Aguirre, José Luís
1978 Catálogo sistemático de los peces marinos que penetran en las aguas continentales de México con aspectos zoogeográficos y ecológicos. Dirección General del Instituto de Pesca, Serie Científica 19, México D.F.
Cooke, Richard G.
1984 Birds and men in prehistoric central Panama. En Recent developments in isthmian archaeology, editado por F. Lange, pp. 243-281. British Archaeological Reports, International Series 212, Oxford.
1989 The use of anurans for food by tropical Amerindians: archaeological and ethnographic evidence. ArchaeZoologia 3: 133-142.
1992a Prehistoric nearshore and littoral fishing in the eastern tropical Pacific: an ichthyological evaluation. Journal of world prehistory 6: 1-49.
1992b Preliminary observations on vertebrate food avoidance by the Precolombian Amerinds of Panama, with comments on the relevance of this behaviour to archaeozoology and palaeoenvironmental reconstruction. En Archaeology and environment in Latin America, Editado por O. Ortiz-Troncoso y T. van der Hammen, pp. 59-107. Instituut voor Pre- en Protohistorische Acheologie Albert Egges van Giffen, Universiteit van Amsterdam, Amsterdam.
1996 Aportes preliminares de la arqueozoología y etnología a investigaciones sobre la taxonomía, ecología y zoogeografía de las especies de la familia Ariidae en el Pacifico Oriental Tropical. Cespedesia (Cali, Colombia) 21:33-43.
Cooke, Richard G. y A.J. Ranere
1989 Hunting in prehistoric Panama: a diachronic perspective. En The walking larder: Patterns of domestication, pastoralism and predation, Editado por J. Clutton-Brock, pp. 295-315. Unwin Hyman, Londres.
1992 Human influences on the zoogeography of Panama: an update based on archaeological and ethnohistorical evidence. En Biogeography of Mesoamerica. Proceedings of a symposium (Mérida, Yucatán, México, October 26-30, 1984), Editado por S.P. Darwin y A.L. Welden, pp. 21-58. Special Publication of the Mesoamerican Ecology Institute.
1994 Relación entre recursos pesqueros, geografía y estrategias de subsistencia en dos sitios arqueológicos de diferentes edades en un estuario del Pacífico central de Panamá. Actas del primer congreso sobre la defensa del Patrimonio Nacional, Panamá 2: 68-114.
1999 Precolumbian fishing on the Pacific coast of Panama. En Pacific Latin America in prehistory: the evolution of archaic and formative cultures. Editado por M. Blake, pp. 103-122. Washington State University Press, Pullman.
Cooke, Richard G., D. Piperno, A.J. Ranere, K. Clary, P. Hansell, S. Olson, W. Valerio, y D. Weiland
1985 La influencia de las poblaciones humanas sobre los ambientes terrestres de Panamá entre el 10,000 a.C. y el 500 d.C. En La Agonía de la naturaleza, Editado por S. Heckadon y J. Espinosa, pp. 3-25. IDIAP/STRI, Panamá.
Cooke, Richard G., G. Tapia R.
1994a Marine and freshwater fish amphidromy in a small tropical river on the Pacific coast of Panama: a preliminary evaluation based on gill-net and hook-and-line captures. En Fish exploitation in the past. (Proceedings of the 7th Meeting of the ICAZ Fish Remains Working Group), Editado por W. van Neer, pp.99-106. Annales du Musée Royale de l'Afrique Centrale, Sciences Zooologiques 274.
1994b Stationary intertidal fish traps in estuarine inlets on the Pacific coast of Panama: descriptions, evaluations of early dry season catches and relevance to the interpretation of dietary archaeofaunas. Proceedings of meeting of fish working group of the international council for zooarchaeology. Offa 51: 287-298.
Day, J.W., Jr., Hall, C.A.S., Kemp, W.M., and Yáñez-Arancibia, A.
1989 Estuarine ecology. Wiley, New York
D'Croz, L., Rivera, R. y Pineda, E.
1977 Observaciones sobre un arte de pesca fija en las costas de la Bahía de Panamá. Conciencia: 14-17.
Kerbis, J.C.
1980 The analysis of faunal remains from the Vidor site. Vínculos (Costa Rica) 6: 125-140.
Vásquez, R. y Thorson, T.B.
1982 The bull shark (Carcharhinus leucas) and largetooth sawfish (Pristis perotteti) in Lake Bayano, a tropical man-made impoundment in Panama. Environmental biology of fish 7: 341-347.
NOTAS
* Este artículo fue publicado originalmente en la revista Arqueología de Panamá La Vieja, avances de investigación, Época Colonial, Rovira y Martín Eds. Patronato Panamá Viejo, agosto 2001.
** Laboratorio de Arqueozoología, Instituto Smithsonian de Investigaciones Tropicales, Ancón, Panamá.
*** Instituto Smithsonian de Investigaciones Tropicales, Ancón, Panamá. E-mail: cooker@naos.si.edu o cominata@hotmail.com
[2] Para información específica del pozo ver Brizuela y Mendizábal en Arqueología de Panamá La Vieja Avances de investigación Época colonial, 2001.
[3] Tres hiomándibulas derechas arrojan un NMI-1de 3. Aunque una muestra que contiene una hiomandíbula derecha, un opérculo izquierdo y una vértebra cervical supone un NMI-1 de 1, si un análisis comparativo o alométrico demuestra que cada elemento anatómico pertenece a un individuo diferente, el NMI se eleva a 3. Por lógica, cuanto más varían los tamaños de los individuos, tanto más fácil es estimar el NMI-2.
[4] El esquema presentado en el Cuadro 6 obedece a observaciones hechas por John Bort y Richard Cooke durante los últimos veinte años sobre la distribución de las especies de peces en la Bahía de Parita respaldadas por los resultados de un proyecto de pesca experimental en el río Santa María y de >80 colectas hechas en una trampa estacionaria o atajo erigido en un estero cerca de Aguadulce (Cooke y Tapia 1994a, 1994b). En vista de que cada estuario en el trópico tiene sus propias características físicas y zoogeográficas, sería temerario suponer que las especies de peces representadas en esta muestra se portarían forzosamente de igual manera en las zonas aledañas a Panamá La Vieja.
[5] Es interesante que los pescadores de Aguadulce llaman a esta especie cominate de Golfo y a Arius platypogon, congo del Golfo en honor a su preferencia por aguas relativamente profundas a cierta distancia de la costa.
Buscar en esta seccion :